 This page explores in greater depth some of the ideas introduced in my article in the May 2020 edition of The Field magazine.
It is available to read online
This page explores in greater depth some of the ideas introduced in my article in the May 2020 edition of The Field magazine.
It is available to read online
A companion article How does a trout catch a fly? was written during the first COVID-19 lockdown.
Some words of explanation. This is a long page. The contents cover topics some of which will be familiar to anglers, other to scientists working in various areas. The aim is to explore the possibility that a Recognition Heuristic controls an element of trout behaviour - the rise to an artificial trout fly. I have tried to make the material accessible, acceptable & hopefully interesting to both audiences, but recognize that some of it may cause 'eyes to glaze' over!
Summary
"Why does a trout take your fly?" - Because it's recognized
The links in this Summary take you to sections on this page that discuss topics in depth.
This essay begins with the views of an American, Thaddeus Norris (1811-1877), who anticipated in 1865 some of the problems we face today with the continued dominance of prescriptive elements of a fly-fishing theory developed by Alfred Ronalds (1802-1860) and Frederic M. Halford (1844 – 1914).
 Halford's dry-fly modus operandi encountered several criticisms over the years. For example:
Halford's dry-fly modus operandi encountered several criticisms over the years. For example:
I agree with Bob Wyatt that we should favour 'simplicity' and, where possible, a science-based answer to the question "Why does a trout take my fly?"
 The underlying simplicity of many animal behaviours has been remarked upon by scientists and fly-fishers:
The underlying simplicity of many animal behaviours has been remarked upon by scientists and fly-fishers:
My simple answer to the question is that a trout takes a fly because it is recognized as food. The 'prey image' in your artificial fly needs to match the 'search image' used by a trout searching for food. That's the short answer I give when I'm guiding. The angler often wants a straightforward answer without the detailed underlying reasons that are discussed in this article. My approach is a 'heuristic' way of avoiding giving a mind-numbing exposition to someone who just wants to go fishing.
What is a heuristic? It's a rule-of-thumb, a simple way that humans, animals, organizations and machines use to quickly make decisions, and find solutions to complex problems. Some psychologists I worked with tended to view human decision-makers as possessing supernatural powers of reason, limitless knowledge, and endless time ( Todd and Gigerenzer 2000). You might get the same impression about trout when reading some fly-fishing literature.
 Trout need to make decisions quickly on the basis of very little information, especially in the 'uncertain' environment of fast-flowing freestone rivers. I suspect they do this on the basis of employing selective attention, and a search image, as input to a Recognition Heuristic to accept a fly as food.
Trout need to make decisions quickly on the basis of very little information, especially in the 'uncertain' environment of fast-flowing freestone rivers. I suspect they do this on the basis of employing selective attention, and a search image, as input to a Recognition Heuristic to accept a fly as food.
Recently psychologists have begun studying how animals and people use heuristics to make decisions on the basis of scant details. And their findings are surprising.
In this long video Gerd Gigerenzer describes the use of heuristics by humans and animals to cope in 'uncertain' environments. I have set the video to start about 17 minutes into his talk where he discusses the gaze heuristic that I used as the basis for my essay "How does a trout catch a fly".
Trout live in an 'uncertain' environment; their food is brought to them by the force of rapidly changing currents. Bankers also live in 'uncertain' times, the arrival of the next financial disaster is unpredictable. I found this talk by the banker Andrew Haldane was a useful introduction to the application of heuristics in situations that lack certainty. Trout use 'fixed action patterns' - innate behaviours modified by age and experience - to survive uncertainty. Bankers rely on regulations - the Halfordian approach.
In some situations less knowledge is better than more knowledge for making accurate decisions. This is an example of using a 'recognition heuristic'. (Goldstein & Gigerenzer 2002, Gigerenzer and Goldstein 2011)
The recognition heuristic uses simple rules for searching, stopping the search, and decision-making.
Therefore trout using a recognition heuristic do not need to engage in long 'inspections' of a fly to confirm their decision to eat it.
The recognition heuristic is ideally suited for making decisions in a fast-moving environment, e.g. a trout using a search image to find food. If an object is recognized, then approach and consume it.
How are 'food' and 'non-food' recognized. The distinction between what to eat and what to avoid is the result of experience (learning), or trout are born with that (innate) ability. Goldstein & Gigerenzer (2002) express it thus: "The direction of the correlation between recognition and the criterion can be learned from experience, or it can be genetically coded. "
These effects include:
Trout using a heuristic to recognize prey may do so on the basis of one feature. Gigerenzer & Gray, (2017) express it thus: "Like the gaze heuristic, the recognition heuristic pays attention to only one variable — one which .. animals have discovered through experience and have used to successfully deal with .. food choice .. " [emphasis added] (Gigerenzer & Gray 2017). The gaze heuristic they refer to utilizes a constant angle of approach (one variable) to intercept prey.
(The gaze heuristic is the basis of a companion article How does a trout catch a fly? that was written during the first COVID-19 lockdown.)
What single feature of an artificial fly is used by trout in their recognition heuristic? There is no cut-and-dried answer to that question. The results of scientific research and accumulated fly-fishing wisdom suggest that movement (or lack of it), size, shape, contrast between the fly and its background (contrast polarity) as well as position in the water column are important.
 But it may only requires one of these features to be present to satisfy the recognition heuristic. A trout using a recognition heuristic may not check for the presence or absence of other features.
"...no further information is looked up about the recognized object." (Goldstein & Gigerenzer 2002).
This is in stark contrast to the characteristics of educated selective trout described by Swisher and Richards (2018), Supinski (2014), Hayes and Stazicker (2019).
But it may only requires one of these features to be present to satisfy the recognition heuristic. A trout using a recognition heuristic may not check for the presence or absence of other features.
"...no further information is looked up about the recognized object." (Goldstein & Gigerenzer 2002).
This is in stark contrast to the characteristics of educated selective trout described by Swisher and Richards (2018), Supinski (2014), Hayes and Stazicker (2019).
 The solution is not to get anxious that your fly must look - to the human eye - to be a close imitation of the natural insect.
That is a holdover from Halford's era when precise imitation was considered de rigueur.
Look at your flies from a trout's point of view. Trout respond to artificial flies designed to be a 'prey image' that incorporates 'triggers / sign stimuli' that fit their 'search image'. A good place to start is Datus Proper's (1993) book which describes several general purpose flies that present prey images for so-called selective trout. It's also worth following Edward R. Hewitt's advice which is still advocated in the fly-fishing literature: change the size of your fly before considering other possibilities.
The solution is not to get anxious that your fly must look - to the human eye - to be a close imitation of the natural insect.
That is a holdover from Halford's era when precise imitation was considered de rigueur.
Look at your flies from a trout's point of view. Trout respond to artificial flies designed to be a 'prey image' that incorporates 'triggers / sign stimuli' that fit their 'search image'. A good place to start is Datus Proper's (1993) book which describes several general purpose flies that present prey images for so-called selective trout. It's also worth following Edward R. Hewitt's advice which is still advocated in the fly-fishing literature: change the size of your fly before considering other possibilities.
This article uses a case study approach to address the question:
"Why does a trout take your fly?"
In situations where quantitative data is deficient in some way (e.g. lacking, unreliable or disputed), a case study approach (Crowe et al, 2011) may provide useful insight into behaviour(s) that cannot be studied under laboratory conditions.
But it may only requires one of these features to be present to satisfy the recognition heuristic. A trout using a recognition heuristic may not check for the presence or absence of other features.
"...no further information is looked up about the recognized object." (Goldstein & Gigerenzer 2002).
This is in stark contrast to the characteristics of educated selective trout described by Swisher and Richards (2018), Supinski (2014), Hayes and Stazicker (2019).
The solution is not to get anxious that your fly must look - to the human eye - to be a close imitation of the natural insect.
That is a holdover from Halford's era when precise imitation was considered de rigueur.
Look at your flies from a trout's point of view. Trout respond to artificial flies designed to be a 'prey image' that incorporates 'triggers / sign stimuli' that fit their 'search image'. A good place to start is Datus Proper's (1993) book which describes several general purpose flies that present prey images for so-called selective trout. It's also worth following Edward R. Hewitt's advice which is still advocated in the fly-fishing literature: change the size of your fly before considering other possibilities.
The UK and the USA: Two countries united by a common misunderstanding of trout behaviours - Selectivity
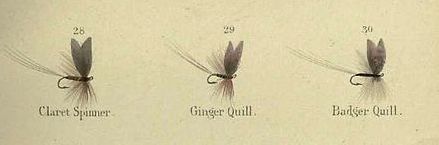 The title refers to the transatlantic sharing of fly-fishing ideas and techniques during the 19th and 20th Century. In the early years the British sent the Americans these dainty high-riding dry flies tied to tempt fastidious English trout.
The title refers to the transatlantic sharing of fly-fishing ideas and techniques during the 19th and 20th Century. In the early years the British sent the Americans these dainty high-riding dry flies tied to tempt fastidious English trout.
 At the end of the century, the Americans gave us low-riding dry flies designed to catch their ultra-selective trout. Our flies didn't really "cut the mustard" over there, but their No-Hackle flies work just fine over here, possibly because they were available here in 1496.
At the end of the century, the Americans gave us low-riding dry flies designed to catch their ultra-selective trout. Our flies didn't really "cut the mustard" over there, but their No-Hackle flies work just fine over here, possibly because they were available here in 1496.
We sent them Halford's dry-fly theory in the hope that “A burden shared is a burden halved.”
This essay focuses on fishing with a dry fly, and introduces several - perhaps lesser known - early American authors who make some valid points for people starting to fish on fast-flowing freestone rivers, such as the East and West Dart, the Devonshire Avon and River Yealm. The challenges faced on these rivers are different from those on chalk streams - the foundry of dry-fly fishing in England.
Thaddeus Norris (1811-1877) and Alfred Ronalds (1802-1860)
 Thaddeus Norris was a 19th century American critic of English flies, the need for strict imitation in artificial flies as well as the emphasis on entomology.
He was probably reacting to a particular chapter in Alfred Ronald's 1836 book The Fly-fisher's Entomology .
Thaddeus Norris was a 19th century American critic of English flies, the need for strict imitation in artificial flies as well as the emphasis on entomology.
He was probably reacting to a particular chapter in Alfred Ronald's 1836 book The Fly-fisher's Entomology .
The real meat of Ronalds' book was Chapter IV: Of a Selection of Insects, and Their Imitations, Used in Fly Fishing. Here, for the first time, the author discussed specific artificial fly imitations by name, associated with the corresponding natural insect. Organized by their month of appearance,
(Wikipedia 2020).
Several eminent fly-fishing historians highlight the impact and continuing importance of Ronalds' contribution to fly-fishing culture (see Wikipedia 2020).
Paul Schullery sums up Ronalds' lasting importance: "Competing theories as entrenched as these do not disappear, and never have, but what did happen both in England and much later in America, was the production of ever more sophisticated tracts on imitation theory"...Ronalds set a course for fly fishing that has never been changed significantly. (Schullery 1987 p85).
It's interested to compare Norris' criticisms written 150 years ago with the current emphasis on 'selectivity' in some British ( Hayes & Stazicker 2019), and American (Swisher and Richards 2018) fly-fishing theory. Selectivity is a veritable oak tree which grew from an acorn planted in 1836 by Ronalds.
Norris tried to fell this mighty English oak when it was a mere sapling.
"It seems to me no more appropriate place than to say a few words about the "routine" and "strict imitation system" which some English writers advocate so stenuously." Norris dismisses the "routine" theory of particular flies for specific months - what we know today as hatch charts - as "pedantic humbug", and, anticipating Parrella (2013) Norris has this to say about entomology: "An extensive knowledge of flies and their names can hardly be of much practical advantage." (Norris 1865 p313.)
 However, Norris accepts the underlying logic of the strict imitation system", but with this thoughtful reservation: "I cannot concede that Trout will rise more readily at the artificial fly which most closely resembles the natural one, for the fish's attention is first attracted because of something lifelike falling on the water, or passing over the surface, and he rises at it because he supposes that it is something he is in the habit of feeding upon, or because it resembles an insect, or looks like a fly, not that it is any particular insect or fly; for we sometimes see the most glaring cheat, which resembles nothing above the waters or below the waters, a piece of red flannel, for instance, or the fin of one of their own species, taken greedily" [
underline added to replace the emphasis in italics in original text]
However, Norris accepts the underlying logic of the strict imitation system", but with this thoughtful reservation: "I cannot concede that Trout will rise more readily at the artificial fly which most closely resembles the natural one, for the fish's attention is first attracted because of something lifelike falling on the water, or passing over the surface, and he rises at it because he supposes that it is something he is in the habit of feeding upon, or because it resembles an insect, or looks like a fly, not that it is any particular insect or fly; for we sometimes see the most glaring cheat, which resembles nothing above the waters or below the waters, a piece of red flannel, for instance, or the fin of one of their own species, taken greedily" [
underline added to replace the emphasis in italics in original text]
(Norris 1865 p333-334. )
 Maybe Norris' views are not influential today (Girard 2003) because he is not read by today's American anglers (Schullery 2007). This also extends to more modern American fly-fishing books. The picture, of a sale of fly-fishing books for $1 each, was taken by John Juracek in his local library in West Yellowstone, Montana (Juracek 2020a).
On sale were books by Lyons and Traver, Brooks and Bergman, LaFontaine and Schwiebert. I've added a link for Travers out of a sense of irony.
Maybe Norris' views are not influential today (Girard 2003) because he is not read by today's American anglers (Schullery 2007). This also extends to more modern American fly-fishing books. The picture, of a sale of fly-fishing books for $1 each, was taken by John Juracek in his local library in West Yellowstone, Montana (Juracek 2020a).
On sale were books by Lyons and Traver, Brooks and Bergman, LaFontaine and Schwiebert. I've added a link for Travers out of a sense of irony.
Nevertheless, I see a lot of Norris' influence lying unnoticed in the corners of contemporary fly-fishing literature. Norris was reporting fly-fishing techniques that were being used - and continue to be used - by what Schullery describes as 'nonimitationists'; anglers without social trappings who were seen as "primitive, unsophisticated, or simply unfashionable" (Schullery 1987 p85).
Schullery points out the paucity of books written by non-imitationists compared to the large imitationist literature. The closest we have to non-imitationists are those who recognize the importance of presentation, and convey a nuanced marriage between presentation and selectivity (Clarke 1996, Juracek et al. 2020c, Rolston 2020, Wyatt 2013). Obviously I don't want to label these people with Schullery's description - some of them are my friends !
If Norris came back to fish today he would understand our concern with effective presentation. He chose to express how a fly should land with this line of poetry: "Light as falls the flaky snow". This delicate presentation can be achieved "if the line is light, the leader fine, and the cast not too long" (Norris 1865 p331).
As a fishing guide there are certain topics I have learnt to avoid with experienced anglers who hold strong views - leader design and length is one of them. I agree with Norris that a transparent leader landing on the water "...does not frighten the fish, but it is the incautious approach or conspicuous position of the angler". Gierach (1989) remarks that 'stealth' is rarely given the prominence it requires in the contemporary fly-fishing literature. Norris writing on careful slow wading gives timeless advice.
Norris wrote something else so important that it requires a separate essay that I hope to write sometime in the future.
"The advice of English writers to fish up stream, or with the wind at one's back, in most cases cannot be followed; for our rough rapid streams in the first instance, and the thickly wooded banks in the other, which make it necessary to wade, ignore both rules. The force of the current in many a good rift would bring the flies back, and, as I have seen with beginners, entangle them in the legs of his pantaloons."
(Norris 1865 p331) [emphasis added]
Norris' insights are all the more remarkable because he was writing before dry-fly fishing was widely practiced in America. He described some anglers false casting to present a dry fly above a 'stretcher' (Norris 1865 p333). This predates Baigent's two-fly technique.
[for description of stretcher = point fly see Gaskell 2019]
There is a lot of common ground shared by Norris, and an emerging understanding of trout behaviours:
Thaddeus Norris was a 19th century American critic of English flies, the need for strict imitation in artificial flies as well as the emphasis on entomology.
He was probably reacting to a particular chapter in Alfred Ronald's 1836 book The Fly-fisher's Entomology .However, Norris accepts the underlying logic of the strict imitation system", but with this thoughtful reservation: "I cannot concede that Trout will rise more readily at the artificial fly which most closely resembles the natural one, for the fish's attention is first attracted because of something lifelike falling on the water, or passing over the surface, and he rises at it because he supposes that it is something he is in the habit of feeding upon, or because it resembles an insect, or looks like a fly, not that it is any particular insect or fly; for we sometimes see the most glaring cheat, which resembles nothing above the waters or below the waters, a piece of red flannel, for instance, or the fin of one of their own species, taken greedily" [
underline added to replace the emphasis in italics in original text] (Norris 1865 p333-334. )
Maybe Norris' views are not influential today (Girard 2003) because he is not read by today's American anglers (Schullery 2007). This also extends to more modern American fly-fishing books. The picture, of a sale of fly-fishing books for $1 each, was taken by John Juracek in his local library in West Yellowstone, Montana (Juracek 2020a).
On sale were books by Lyons and Traver, Brooks and Bergman, LaFontaine and Schwiebert. I've added a link for Travers out of a sense of irony.For me, Norris' great insight is what he says about trout. His focus is on the trout; what the trout sees, and how the trout responds to what it perceives (von Uexkull 1934). He is not concerned with how closely artificial flies look like natural insects to human inspection.
Frederic M. Halford (1844 – 1914)
The use of dry flies before Halford is described in detail by Dr Andrew Herd (2002).
Halford's 1886 book "Floating Flies and How to Dress Them" enjoys an international reputation for promoting a method of catching trout by:
 My impression, reading Halford (1886), is that he is describing a method that tries to give the angler the best chance of catching a rising fish. He is certainly aware of the need for stealth, as well as delicate and accurate presentation (p 123). He advocated upstream casting, but if necessary a cast could be made across stream provided that the tip of the rod was moved downstream to avoid drag. (p124). If necessary, a fly cast downstream can be "efficacious" (p124).
My impression, reading Halford (1886), is that he is describing a method that tries to give the angler the best chance of catching a rising fish. He is certainly aware of the need for stealth, as well as delicate and accurate presentation (p 123). He advocated upstream casting, but if necessary a cast could be made across stream provided that the tip of the rod was moved downstream to avoid drag. (p124). If necessary, a fly cast downstream can be "efficacious" (p124).
In 1886, Halford was careful not place dry-fly fishing on Southern chalk streams above wet-fly fishing on Northern freestone rivers. He deprecated the use of the phrase "chuck and chance it" to stigmatize wet flies that had appeared in The Fishing Gazette (p117). And he expressed similar admiration of the skills of Northern wet-fly anglers in his second book (Halford 1889 p37)
Halford's method is said to have transformed fly-fishing for trout into a scientific endeavour (Gubbins, 2018, Schweibert, 1979). I think this was principally in the area of what is known today as 'matching the hatch', which has the feel of a 'will-o'-the-wisp' venture.
Halford benefited from knowing the fly-fishing entomologist Martin Mosely who worked at The Natural History Museum in London (Barnard 2011). Mosely is credited with attempting to persuade fly-fishers to name artificial flies on the basis of what they represented (their scientific Latin name), rather than when they were effective (e.g. March Brown, February Red), or who inspired their creation (e.g. Greenwell's Glory, Adams). (Parrella 2013).
Apparently, there are 630 species of up-winged flies / mayfly (Ephemeroptera) in North America, but only 51 in the British Isles. And 15,000 artificial flies (Banasch 2000 quoted by Parrella).
Halford (1913 p154-5) appreciated that there are limits to what an angler can confidently know about the insect taken by a trout: "As a rule, one is too far from the rising fish to be able to distinguish with any degree of certainty the genus and species of the insect on which it is feeding." Halford's advice was to avoid changing the artificial fly in this situation, or when there was a 'masking hatch' because it "soon degenerates into hammering, which is an infallible means of educating a trout and making it shy"
 I detect a sharp change of tone in Halford's last book published in 1913. The title of Chapter 3 "The Ethics of the Dry Fly" sets the mood. As a past chairman of a local fishing association, I smelt trouble brewing when I read this comment about the consequences of casting to a fish that wasn't rising:
I detect a sharp change of tone in Halford's last book published in 1913. The title of Chapter 3 "The Ethics of the Dry Fly" sets the mood. As a past chairman of a local fishing association, I smelt trouble brewing when I read this comment about the consequences of casting to a fish that wasn't rising:
"His chance of success is in no case very good, and on club or subscription waters, which are usually very much overfished, he is far more likely to scare the fish than to induce it to rise at his fly. ". The complaint is that this is a wet-fly tactic, and to make matters worse they fish with a cast of 2,3 or even 4 wet flies. And if you still haven't got the message "It is the very worst of taste for a guest to follow these tactics, because he should know that it is the unwritten law of the dry-fly man on a chalk-stream to eschew any but the legitimate method. "
Maintaining a chalk stream is an expensive undertaking even in Halford's time. The British are reluctant to talk about private wealth, and Halford would have believed that a guest should consider the financial burden placed on the host by their visit.
Halford suggests that casting a nymph upstream is just as bad as downstream wet-fly fishing because it targets fish that are not rising, and is a breach of etiquette:
"But in my view the continual flogging and the continual movement of the angler making his way along the bank, too often in full view of the trout, are, however, very nearly as destructive of the confidence of the fish as downstream fishing. "
Andrew Herd (2002) describes Halford's reference to "The Etiquette of Golf" as a way of dealing with anglers who flout "The Ethics of the Dry Fly". But Dr Herd spared his readers mention of the punishment handed out to golfers, and fly-fishers who transgressed, social ostracism: "He will find difficulty in persuading his fellow-members to make a friendly match with him. I would suggest that the ethics of the dry-fly on a fishery where dry-fly only is permitted should be regarded in much the same light. "
 I have detected a tendency to absolve Halford of responsibility for the rancour that arose around dry-fly fishing, and instead focus on the attitude of Halford's followers. I'm not sure. The social ostracism recommended by Halford is a powerful way to control one's fellow beings, for example, "being sent to Coventry" or the treatment received by Captain Boycott pictured.
I have detected a tendency to absolve Halford of responsibility for the rancour that arose around dry-fly fishing, and instead focus on the attitude of Halford's followers. I'm not sure. The social ostracism recommended by Halford is a powerful way to control one's fellow beings, for example, "being sent to Coventry" or the treatment received by Captain Boycott pictured.
To an outsider this appears very snobbish, and it is. In some quarters dry-fly fishing is viewed as 'exclusive' to this day (Cohen 2016), but here is another view with a twist at the end - Tapply (2016). Trout can be unforgiving social-levellers. To be fair to Halford, I think he may have needed to conserve trout stocks in the days before the advent of catch-and-release. Halford was renting his fishing, maintaining the river, and stocking to supplement the natural head of fish (Halford 1913 p74). Under the sub-heading "The degeneration of chalk-stream trout" (p392), Halford (1913) commented: "They do not grow to the dimensions of those we killed in bygone days, they do not rise as freely, they do not play as well, they are not as handsome in shape, colour, or markings, and they are not as palatable on the table." Similar problems were encountered with stocking in North America (Michigan) (Gubbins 2018).
This, Halford's last book, was published a year before he died. He has just described the decay to the central pillar of his approach to fly fishing - stocking with fish that were disinclined to rise. Today, visitors to the Test are keen to catch a trout on a dry fly, but they often have to resort to a nymph - the method introduced by his nemesis, Skues (Cooper and Jardine 2020).
Skues and Halford
 "Remember Skues began his serious fly fishing career under the shadow of Halford and was in fact a follower of the Halford doctrine." (Travis 2016).
"Remember Skues began his serious fly fishing career under the shadow of Halford and was in fact a follower of the Halford doctrine." (Travis 2016).
The first, and still famous, challenge to Halford came from Skues who pointed out that trout could be caught on a nymph presented a few inches below the surface. Unfortunately, this 'minor tactic' generated more heat than the scientific light it deserved. Perhaps in order to gain acceptance from dry-fly purists, Skues retained the essential elements of Halford's theory: he cast upstream to a feeding trout with an artificial fly tied to represent an insect that autopsy confirmed had been consumed shortly before capture. Skues (1914) simply modified Halford's technique by altering the position of the artificial fly in the water column.
Gingrich (1974) suggested that Skues effected a revolution, and Schullery (2008) called Skues a tentative rebel. I don't see Skues in those terms. Skues himself introduced the nymph "to be used as a supplement to, and in no sense to supplant or rival, the beautiful art of which Mr. F. M. Halford is the prophet." (Skues 1914).
From a scientific point of view Skues' ideas are no threat at all to Halford's theory, but it enraged Halford and his disciples. For example, this quote from Halford tells us more about Halford's state of mind than any threat from Skues: "Those of us who will not in any circumstances cast except over rising fish are sometimes called ultra purists and those who occasionally will try to tempt a fish in position but not actually rising are termed purists... and I would urge every dry fly fisher to follow the example of these purists and ultra purists." (Halford 1913 p 69-70, and quoted by Bark (1992) in " A History of Flyfishing")
Skues borrowed heavily from Halford to present a parallel method of catching trout. The heat generated by the debate in the Flyfishers' Club in 1938 is of interest to students of minor human interactions, but the debate itself failed to illuminate the merits of complementary approaches that exploit the reaction of trout to food on (Halford) or below (Skues) the water surface.
 To avoid
'confirmation bias', I read the report of "The Nymph Debate" in The Fly fishers Club Spring Journal (1938), before reading Lawton's (2005) assessment.
We came off lightly down here in the West Country.
The Rev. Percy Sheriffs called for "a plague on both your houses", and a ban on nymphs and dry flies to conserve trout stocks !
To avoid
'confirmation bias', I read the report of "The Nymph Debate" in The Fly fishers Club Spring Journal (1938), before reading Lawton's (2005) assessment.
We came off lightly down here in the West Country.
The Rev. Percy Sheriffs called for "a plague on both your houses", and a ban on nymphs and dry flies to conserve trout stocks !
I found the account of the debate depressingly familiar, and agree with Lawton who found "..the argument against nymph fishing was based on ignorance, intolerance and prejudice. It was ever thus."
Perhaps, sadly, the lasting legacy of Halford's 1913 book is responsibility "for a kind of sporting elitism that still dogs the dry fly" (Herd 2002, Gubbins 2018). And the debate continues to this day - on English chalk streams ... (Cooper & Jardine 2020).
Personally I don't detect a unbridgeable chasm between Halford and Skues. Both advocated casting upstream to trout seen rising to an identified insect. In 1921, Skues discovered a tool that enabled him to identify what the trout were feeding on - a marrowspoon to extract the contents of a trout’s stomach for examination on a white china plate. This overcomes a major obstacle to Halford's approach - waiting to see exactly what dun a trout was taking - searching for a 'will-o'-the-wisp' which sometimes involves a long wait.
Replication and extension: Doubts from abroad
For a scientist it is important that observations can be replicated by independent laboratories. And that is just what happened when Halford was approached by two Americans: Theodore Gordon and George La Branche for advice on dry-fly fishing.
Theodore Gordon
Theodore Gordon received flies, and corresponded with Halford and Skues, but needed to alter the English flies to match the insects in the Catskills region (Schullery 2002). This was to be expected; insects vary between regions and this alone does not challenge Halford's theory of precise imitation.
George La Branche: A fresh pair of eyes and a logical mind
 In 1914 George La Branche published The Dry Fly and Fast Water. La Branche had spotted a logical flaw in Halford's theory. He wrote: When fish are feeding upon some particular species of insects it is quite logical to assume that an imitation of that species will appeal to them more readily than an imitation of any other. But when insects are numerous, as they are on occassions, and the fish are moving about, the chance of the artificial fly being selected from among the great number of naturals on the water is one to whatever the number may be. (La Branche 1914 p60 emphasis added).
In 1914 George La Branche published The Dry Fly and Fast Water. La Branche had spotted a logical flaw in Halford's theory. He wrote: When fish are feeding upon some particular species of insects it is quite logical to assume that an imitation of that species will appeal to them more readily than an imitation of any other. But when insects are numerous, as they are on occassions, and the fish are moving about, the chance of the artificial fly being selected from among the great number of naturals on the water is one to whatever the number may be. (La Branche 1914 p60 emphasis added).
The logical problem - of a trout selecting an imitation fly amongst a large hatch of identical insects (selectivity) - spotted by La Branche exists to this day on rivers with substantial fly hatches.
Furthermore, in contrast to Halford and dry-fly purists, La Branche advocated fishing the water rather than casting to a specific rising trout. This was necessary because of the river conditions he faced - few rising trout and fast water. These conditions will be familiar to fly-fishers on Dartmoor and other spate rivers.
La Branche did not consider himself to be a dry-fly purist. He once remarked:
 I consider that the real purist wastes countless joyous and active hours waiting, according to theory, for the fish to rise before he starts. That was standard behaviour in England. La Branche, not one to waste minutes, let alone hours, learned to fish dry flies in fast water, casting to trout he knew were there. but which had not revealed themselves by rising. (Betts 2002)
I consider that the real purist wastes countless joyous and active hours waiting, according to theory, for the fish to rise before he starts. That was standard behaviour in England. La Branche, not one to waste minutes, let alone hours, learned to fish dry flies in fast water, casting to trout he knew were there. but which had not revealed themselves by rising. (Betts 2002)
La Branche challenged Halford's theory in another fundamental way that remains a live issue to this day.
La Branche rejected Halford's insistence on using a precise imitation of the natural fly. Today we use the term selectivity to refer to this central tenet of Halford's theory.
La Branche quotes, with approval, Professor of Zoology James Rennie who wrote in 1833: I have used the phrase "pretended imitation" as strictly applicable to by far the greater number of what are called by anglers artificial flies, because these rarely indeed bear the most distant resemblance to any living fly or insect whatever, though, if exact imitation were an object, there can be little doubt that it could be accomplished much more perfectly than is ever done in any of the numerous artificial flies made by the best artists in that line of work. (Rennie 1833, p 137-8)
 La Branche's attitude prompted a debate with Louis Rhead in the 1920s on exact imitation in tying trout flies. It was held at The Anglers' Club of New York. The confrontation was described in a letter written by Eugene V. Connett 111. Louis Rhead is described as, a conceited little fellow, a very delightful artist, and the worst fly designer that God ever put breath in. La Branche chewed poor Rhead into small pieces and spit them out. It was quite cruel and I always felt a bit guilty about having arranged the debate:' Connett wrote. (Belknap, 1992)
La Branche's attitude prompted a debate with Louis Rhead in the 1920s on exact imitation in tying trout flies. It was held at The Anglers' Club of New York. The confrontation was described in a letter written by Eugene V. Connett 111. Louis Rhead is described as, a conceited little fellow, a very delightful artist, and the worst fly designer that God ever put breath in. La Branche chewed poor Rhead into small pieces and spit them out. It was quite cruel and I always felt a bit guilty about having arranged the debate:' Connett wrote. (Belknap, 1992)
 La Branche's deviation from Halford did not go unnoticed by Theodore Gordon. Paul Schullery (1987 p119-120) refers to a letter donated to the American Museum of Fly Fishing by La Branche's daughter written by her father probably in the 1950s in which he describes Gordon as ... A great friend and companion. But then reveals that Gordon ... told me that I was ?belittling? (word unclear) the theory of dry fly fishing. He (Gordon) agreed with Dewar and Halford that what I was doing was an affectation and that the dry fly should be used on slow flowing water over rising fish only. I (LaBranche) was upset more than a little, but persevered with my idea.
La Branche's deviation from Halford did not go unnoticed by Theodore Gordon. Paul Schullery (1987 p119-120) refers to a letter donated to the American Museum of Fly Fishing by La Branche's daughter written by her father probably in the 1950s in which he describes Gordon as ... A great friend and companion. But then reveals that Gordon ... told me that I was ?belittling? (word unclear) the theory of dry fly fishing. He (Gordon) agreed with Dewar and Halford that what I was doing was an affectation and that the dry fly should be used on slow flowing water over rising fish only. I (LaBranche) was upset more than a little, but persevered with my idea.
Baigent: An independent Yorkshire voice
Norris and La Branche were not alone in finding that the approach - developed in relatively sedate English chalk streams - was not an applicable technique in faster-flowing freestone (spate) rivers.
 La Branche corresponded with Dr William Baigent (1862-1935) a medical doctor in Northallerton, a market town in North Yorkshire (UK). He is remembered for Baigent’s Brown, and his influence on the Catskill tradition of fly-tying through correspondance with American authors George La Branch and Preston Jennings. (Rob Smith, 2018).
La Branche corresponded with Dr William Baigent (1862-1935) a medical doctor in Northallerton, a market town in North Yorkshire (UK). He is remembered for Baigent’s Brown, and his influence on the Catskill tradition of fly-tying through correspondance with American authors George La Branch and Preston Jennings. (Rob Smith, 2018).
Like La Branche , Baigent had struck out on his own. He did not follow the local fly-fishing culture. Born and living his life in North Yorkshire he fishing dry flies in an area of England where fishing soft-hackled wet flies reigned supreme. He fished dry flies at a time when the Halfordian doctrine of precise imitation was de rigueur. He eschewed precise imitation, instead devoting many years to breed Old English Game Cock to supply hackle to tie his unique 'variant' style of flies including this long-hackled Baigent's Brown. Asked what the Baigent Brown was tied to represent, Baigent replied: "It is not tied to represent any fly, it is tied to catch trout.(Roberts, 1994, p 215)
 The influence of Baigent is clear in the four dry flies in the corners of this picture of a collection of La Branche's flies held by the American Museum of Fly Fishing (Bett, 2002)
The influence of Baigent is clear in the four dry flies in the corners of this picture of a collection of La Branche's flies held by the American Museum of Fly Fishing (Bett, 2002)
It's surprising that La Branche did not adopt Baigent's two-fly technique: If trout are nymphing, a nymph or wet fly could be mounted on the point, whilst a dry fly could be mounted on the dropper. Nicholas Fitton (1992) gives a detailed description of the history, and his experience using this almost-forgotten technique which is very effective on local rivers whether or not fish are rising.
Baigent's influence on American fly designs live on; Swisher and Richards (1975 p.72) recommended searching the water with "heavily hackled" spiders, variants and bivisibles. Proper (1993 p 163) remarks favourably on Baigent's interest in designing flies, rather than copying handed-down patterns.
The Gold Ribbed Hare's Ear: Halford's Achilles' Heel ?
 In Halford's time the Gold Ribbed Hare's Ear (GRHE) was, and continues to be, a very effective pattern. It doesn't look like any natural fly. Halford added wings to make it acceptable to dry fly purists, without any deterioration in its attractiveness to trout. In this comment from his 1886 book Halford recognized its effectiveness.
In Halford's time the Gold Ribbed Hare's Ear (GRHE) was, and continues to be, a very effective pattern. It doesn't look like any natural fly. Halford added wings to make it acceptable to dry fly purists, without any deterioration in its attractiveness to trout. In this comment from his 1886 book Halford recognized its effectiveness.
Perhaps a little cruelly, Skues (1921 p91) wrote: At one time the late Mr. F.M. Halford was a great advocate of the Gold-ribbed Hare's Ear [GRHE], but I believe that latterly his enthusiasm for precise imitation induced him to give it up, successful pattern though he knew it to be, because he could not explain its success to his satisfaction.
 Was Skues being mischievous when he added: "Still, the Hare's Ear kills. And I should like to know who was the genius who first conceived its possibilities, and how he got at his theory. If we had that information progress might be possible " (Skues 1921 p91) [emphasis added].
Was Skues being mischievous when he added: "Still, the Hare's Ear kills. And I should like to know who was the genius who first conceived its possibilities, and how he got at his theory. If we had that information progress might be possible " (Skues 1921 p91) [emphasis added].
In 1889 Halford appreciated that the GRHE was very effective when trout were taking nymphs just below the surface (bulging):
"This pattern is placed first of the series
[Group I.— Imitations Of Natural Insects.
Section I, — Olive Duns]
as the most successful of modern times. From early spring to late autumn it is one of the most killing of all the duns, and is, besides, pre- eminently the fly to be recommended for bulging or tailing fish. It is probably taken for the sub-imago emerging from the larval envelope of the nymph just risen to the surface." (Halford 1889 p147-8)
 Halford discarded the GRHE because it simply did not fit into his theory that dry-fly fishing required presenting a fly tied to imitate an identified floating dun. Gary La Fontaine recognised that the GRHE forecast Swisher and Richards' No-Hackle dry flies: ".. In a pique of intellectual integrity or foolhardy sophistry, depending on one's point of view, he abandoned the fly because this ancient no-hackle did not 'cock' like a proper dry fly." (La Fontaine 1990 p253)
Halford discarded the GRHE because it simply did not fit into his theory that dry-fly fishing required presenting a fly tied to imitate an identified floating dun. Gary La Fontaine recognised that the GRHE forecast Swisher and Richards' No-Hackle dry flies: ".. In a pique of intellectual integrity or foolhardy sophistry, depending on one's point of view, he abandoned the fly because this ancient no-hackle did not 'cock' like a proper dry fly." (La Fontaine 1990 p253)
This is not the last word about the GRHE. In my opinion, the GRHE may qualify as the prototype for a Supernormal stimulus
The myth of the educated selective trout?
 Like La Branche before him, Marinaro (1970 p 62) was a critic of Halford's strict insistence on precise imitation "Every detail of the natural fly's anatomy must be included, no matter how absurd the result. For example "... the segments of the natural fly were counted and the same number of turns of ribbing were included in the artificial"
Like La Branche before him, Marinaro (1970 p 62) was a critic of Halford's strict insistence on precise imitation "Every detail of the natural fly's anatomy must be included, no matter how absurd the result. For example "... the segments of the natural fly were counted and the same number of turns of ribbing were included in the artificial"
Marinaro uses a strong word, 'absurd', to describe this "out of many, one" approach E pluribus unum . The angler is casting an artificial fly during a hatch of naturals - the statistical odds are stacked against the angler.
Nevertheless Halford's central tenet - using an artificial fly to imitate the natural fly that a trout is feeding on - lives on in concepts such as 'selectivity', 'the highly educated trout' (Dewar 1910 p26) and 'matching the hatch'.
 Originally published in 1971 as Selective Trout , and republished in 2018 as Selective Trout The Last Word on Stream Entomology and Aquatic Insect Imitation, the front cover proclaims that Doug Swisher and Carl Richards have written The book that changed fly fishing in America.
Originally published in 1971 as Selective Trout , and republished in 2018 as Selective Trout The Last Word on Stream Entomology and Aquatic Insect Imitation, the front cover proclaims that Doug Swisher and Carl Richards have written The book that changed fly fishing in America.
We don't need to take the publisher's word for it, America's pre-eminent angling historian Paul Schullery (2006 p 105) judges it: "probably the most far-reaching of all American fly-fishing books since World War II, Doug Swisher and Carl Richards's 'Selective Trout' (1971) elevated our thinking not only in fly-tying theory, but also, through its wonderful photographs of insects, in our basic understanding in what the flies really imitated" [emphasis added].
 This passage from Mason's book, first published in 1988 and republished in 1994, confirms that Selective Trout caused a paradigm shift in American fly-fishing.
This passage from Mason's book, first published in 1988 and republished in 1994, confirms that Selective Trout caused a paradigm shift in American fly-fishing.
The criticism that no-hackle flies were available several centuries ago fades before the central concepts in the book: 'selectivity', 'educated trout', and 'matching the hatch'.
 Wyatt (2013) commented: "Selective Trout, possibly the largest selling fly-fishing book of all time, directly and indirectly provides the theoretical background for much of what has been written since."
Wyatt (2013) commented: "Selective Trout, possibly the largest selling fly-fishing book of all time, directly and indirectly provides the theoretical background for much of what has been written since."
Wyatt wrote What Trout Want: The Educated Trout and Other Myths in response to Selective Trout .
You don't have to read beyond the dust covers to know the authors opinions: Doug Swisher and Carl Richards have written the last words on aquatic imitation; Bob Wyatt regards 'selectivity', 'matching the hatch' and 'the educated trout' as myths. I detect a growing scepticism in sticking too closely to hatch-matching (Swentosky 2019 & comments; Goetz 2019; Clement 2019).
Matching the hatch rests on an unstable foundation.
"This historical lack of rigor in identifying and naming the arthropods on which the sport is based truly confounds the already difficult concept of "matching the hatch" - a paradigm well entrenched in the fly-fishing psyche." (Parrella 2013)
 The still image is taken from Doug Swisher's video Strategies for Selective Trout in which he describes the characteristics of the right fly as being : size, shape and colour in that order.
The still image is taken from Doug Swisher's video Strategies for Selective Trout in which he describes the characteristics of the right fly as being : size, shape and colour in that order.
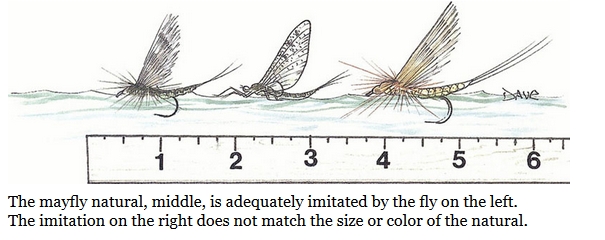 This illustration from Swisher and Richards' book Selective Trout (2018) does seem to perpetuate Halford's attitude to fishing with a dry fly that is a close imitation of the natural. They explain their attitude thus:
This illustration from Swisher and Richards' book Selective Trout (2018) does seem to perpetuate Halford's attitude to fishing with a dry fly that is a close imitation of the natural. They explain their attitude thus:
La Branche corresponded with Dr William Baigent (1862-1935) a medical doctor in Northallerton, a market town in North Yorkshire (UK). He is remembered for Baigent’s Brown, and his influence on the Catskill tradition of fly-tying through correspondance with American authors George La Branch and Preston Jennings. (Rob Smith, 2018).
The influence of Baigent is clear in the four dry flies in the corners of this picture of a collection of La Branche's flies held by the American Museum of Fly Fishing (Bett, 2002)
In Halford's time the Gold Ribbed Hare's Ear (GRHE) was, and continues to be, a very effective pattern. It doesn't look like any natural fly. Halford added wings to make it acceptable to dry fly purists, without any deterioration in its attractiveness to trout. In this comment from his 1886 book Halford recognized its effectiveness.
Perhaps a little cruelly, Skues (1921 p91) wrote: At one time the late Mr. F.M. Halford was a great advocate of the Gold-ribbed Hare's Ear [GRHE], but I believe that latterly his enthusiasm for precise imitation induced him to give it up, successful pattern though he knew it to be, because he could not explain its success to his satisfaction.
Was Skues being mischievous when he added: "Still, the Hare's Ear kills. And I should like to know who was the genius who first conceived its possibilities, and how he got at his theory. If we had that information progress might be possible " (Skues 1921 p91) [emphasis added].
Halford discarded the GRHE because it simply did not fit into his theory that dry-fly fishing required presenting a fly tied to imitate an identified floating dun. Gary La Fontaine recognised that the GRHE forecast Swisher and Richards' No-Hackle dry flies: ".. In a pique of intellectual integrity or foolhardy sophistry, depending on one's point of view, he abandoned the fly because this ancient no-hackle did not 'cock' like a proper dry fly." (La Fontaine 1990 p253)
Like La Branche before him, Marinaro (1970 p 62) was a critic of Halford's strict insistence on precise imitation "Every detail of the natural fly's anatomy must be included, no matter how absurd the result. For example "... the segments of the natural fly were counted and the same number of turns of ribbing were included in the artificial"
Originally published in 1971 as Selective Trout , and republished in 2018 as Selective Trout The Last Word on Stream Entomology and Aquatic Insect Imitation, the front cover proclaims that Doug Swisher and Carl Richards have written The book that changed fly fishing in America.
This passage from Mason's book, first published in 1988 and republished in 1994, confirms that Selective Trout caused a paradigm shift in American fly-fishing.
Wyatt (2013) commented: "Selective Trout, possibly the largest selling fly-fishing book of all time, directly and indirectly provides the theoretical background for much of what has been written since."
The still image is taken from Doug Swisher's video Strategies for Selective Trout in which he describes the characteristics of the right fly as being : size, shape and colour in that order.
This illustration from Swisher and Richards' book Selective Trout (2018) does seem to perpetuate Halford's attitude to fishing with a dry fly that is a close imitation of the natural. They explain their attitude thus:
The fast flowing spate (freestone) rivers we fish run off Dartmoor; they are not the main focus of Swisher and Richards' book: 'The types of streams that are conducive to the dry fly and selective rising trout are:
Therefore you might wonder if Swisher and Richards have any relevance for fly fishing on freestone rivers. Yes they do, under circumstances that we sometimes encounter on local rivers: "There are periods, though, especially during low water in quiet pools, when delicate casting and realistic imitations are needed." [emphasis added] (Swisher & Richards, 1975)
We saw above that La Branche was criticised for dry-fly fishing in rapid water :He (Gordon) agreed with Dewar and Halford that what I (La Branche) was doing was an affectation and that the dry fly should be used on slow flowing water over rising fish only. (Schullery, 1987)
Swisher and Richards explain that trout in slow flowing water become ultra-selective because in placid fertile rivers they enjoy long inspection times that enable them to discriminate between small insects and flotsam drifting towards them. To further complicate dry-fly fishing on these rivers, smaller insects hatch in greater numbers than larger flies, and trout become more selective as the size of the natural insect decreases. "When fishing a #28 (i.e. hook size=28) hatch, for example, a 1-mm variation from the natural means at least a 30 per cent dimensional error - which is disastrous and results in nothing but refusals from the trout." (Chapter 3).
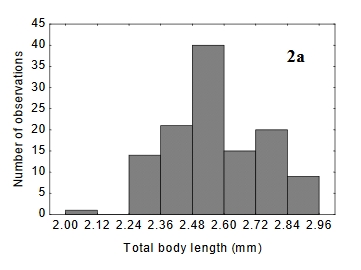 It is very difficult to arrive at any firm conclusion about the impact of "dimensional error" on trout behaviour.
Estimating the body size frequency distribution to determine the average length and size range of particular insects is far from straightforward (Gouws et al. 2011).
It is very difficult to arrive at any firm conclusion about the impact of "dimensional error" on trout behaviour.
Estimating the body size frequency distribution to determine the average length and size range of particular insects is far from straightforward (Gouws et al. 2011).
Gouws (2007 p5-12) reviews the factors that need to be considered. This diagram shows the wide variation in body length in one genus (Nysius ) from a total of 16 different species she measured. (Figure 2a, Gouws 2007 p38).
 An important feature of the first edition (1971) of Selective Trout was the re-introduction of No-Hackle flies with a body that rests in the surface film.
An important feature of the first edition (1971) of Selective Trout was the re-introduction of No-Hackle flies with a body that rests in the surface film.
This represented a significant departure from the conventional Halfordian dry fly at that time tied with hackles wound around the hook so that the fly would land with just hackle and tail touching the water.
One of the flies illustrated here (from the AMFF Dame Juliana Berners Collection ) is tied according to instructions in a book published in 1496 by Dame Juliana Berners. I'll leave you to decide which is which.
Then it's worth taking a few minutes to scroll through this collection of 15th Century flies. They all look like No-Hackle flies to me. They would not have been regarded by Halford as dry flies. In the late 19th and early 20th centuries: "A dry fly is a fly .. which sits with the majority of the fly above the surface of the water" (Herd 2003 p273)
 Schullery (1987, p 101-2) considers that Marinaro's redefinition of dry flies is far more useful than that narrower Halfordian concept: "We must begin with the proposition that no matter how dry the fly is, it must touch the water and be exposed to the air at the same time. If this idea is carried out to its logical conclusion, all of us must agree that if the smallest portion is exposed to the air no matter how deeply submerged the fly may be, it is still a legitimate form of the dry fly," (Marinaro 1970). I had to smile when I realized that Marinaro's fly [illustrated on left] would have passed Halford's 'majority' rule.
Schullery (1987, p 101-2) considers that Marinaro's redefinition of dry flies is far more useful than that narrower Halfordian concept: "We must begin with the proposition that no matter how dry the fly is, it must touch the water and be exposed to the air at the same time. If this idea is carried out to its logical conclusion, all of us must agree that if the smallest portion is exposed to the air no matter how deeply submerged the fly may be, it is still a legitimate form of the dry fly," (Marinaro 1970). I had to smile when I realized that Marinaro's fly [illustrated on left] would have passed Halford's 'majority' rule.
In fairness to Halford, he wrote (Chapter 11 Floating Flies and Sunk Flies) that the floating or dry fly: "... is an imitation of a winged insect, either emerging or emerged from the nymph state, on the surface of the stream..." (Halford 1889 p36) [emphasis added]. Halford was making a sharp distinction between dry and wet fly - a fly presented clear of, and beneath the surface.
You might think that a definition of what constitutes a dry fly is just "dancing on the head of a pin". But it has the practical effect of extending dry flies to encompass 'emergers' and 'cripples' (partially emerged insects) that are represented by No-Hackle patterns.
The No-Hackle approach was based on Vince Marinaro's observation that the wings and body of the natural fly trigger the rise. However
Swisher and Richards No-Hackle dry flies proved unpopular because, as Mike Lawson shows in this video, the wings were difficult to tie in the correct position on the body, and lacked durability. Consequently they fell out of favour.
AK Best relates a revealing anecdote about Swisher and Richards No-Hackle dry flies in the last 5 minutes of this podcast interview . Mike Lawson is recognised as one of the few tyers who can tie perfect wings on No Hackle dry flies. AK Best watched him tie one, and asked: "When you fish that fly what do you do?" Best reported that Lawson "took his thumbnail, and he broke that wing all apart," and said: Now I've got fibres down in the water simulating legs, and there's enough sticking up simulating the wings. Lawson explained "I destroy the wing before I fish it !". AK Best commented that "What sells that fly is a perfect wing profile".

 But the No-Hackle design remains popular because of the effectiveness of the design concept.
We now have a choice of no-hackle flies with - "big-wing-and-soggy-bottom" - features that trigger a trout's rise (Kenyon 2020).
For example,
But the No-Hackle design remains popular because of the effectiveness of the design concept.
We now have a choice of no-hackle flies with - "big-wing-and-soggy-bottom" - features that trigger a trout's rise (Kenyon 2020).
For example,
Local rod builder Luke Bannister using a Sparkle Dun to good effect
Devon based author, and Wild Trout Trust founder, Mike Weaver introduced UK anglers to comparaduns (e.g. Sparkle Dun) in his widely acclaimed 1991 book "The Pursuit of Wild Trout". He continues to demonstrate the tying technique at the annual Snowbee Open Day each Spring. Mike fished with American angler Sid Neff (Agro 2000) over many years - "who has influenced my fly fishing for river trout more than any other angler" (Weaver, 1991, p 5). Their friendship enabled an influential positive transatlantic transfer of fly-fishing innovations - such as the use of deer hair - to the benefit of anglers fishing UK freestone (spate) rivers.

Trigger #1 ...

or Trigger #2 ...

or Trigger #3
The problem faced by trout in these British and American spate (freestone) rivers, are summed up in this passage from John Bett's (2002) essay on La Branche. For a trout in six inches of water, the window above him is twelve inches across. Slow water moves at one foot or less per second; fast water at about four feet or more in the same period of time. In the swifter currents a fly will cross the front half (six inches) of the window in one-eighth of a second or less, depending on the speed of the water. Even for an animal conditioned to these circumstances over millions of years, a tiny fraction of a second is not much time to go through everything needed to discriminate and decide whether to intercept or refuse an object. So the trout has to make the best guess he can.
In summary, No-Hackle flies were invented to cope with selective trout on slow-moving limestone / chalk streams (UK) rivers. Their design was influenced by Marinaro's insights into factors that trigger the rise. No-Hackle flies were found to be effective on both types of river - fast and slow moving - whereas Halford's precise imitation hackled dry flies are relatively ineffective on fast-flowing freestone rivers.
Is selectivity still alive and well ?
I had a pretty relaxed attitude to selectivity until recently. It was probably due to reading too much American fly-tying literature. For example, no one could accuse Dave Hughes of being an utra-selectionist zealot: "It's another minor axiom of this book that if something resembles something else so closely that anglers and trout mistake it for whatever else it looks like, then a pattern that illustrates the something else will work fine, and there's no need to create and carry a new one. " (Hughes 2005). Hughes manages to absorb, without undue fuss, Swisher and Richards emphasis on selectivity - into a book that caters for fly-fishing on a variety of river types - by introducing the term 'habitat' into the factors to take into account when selecting a suitable fly.
But when I probed deeper I realized that I shouldn't jump to conclusions. For example, fly-fishing author with a degree in Fisheries Biology, John Juracek wrote about angler's search for a fly to meet the demands of selective trout:
Certain beliefs are so widely and deeply entrenched in our sport that they’re essentially considered givens, and rarely, if ever, called into question. One of the most closely held says that fly pattern matters. Matters in terms of success. We’re taught to believe that our choice of fly is responsible for the fish we catch, and that if we merely find the right fly, our success will know no bounds. But does it really work that way? Is our choice of fly that critical? Good question. (John Juracek (2016) )
I detect two forms of selectivity in the UK fly-fishing literature: 'selective trout' / 'matching-the-hatch' and 'selective-educated trout'. The more reasonable form is expressed by Pat O'Reilly in the Preface to his excellent book "Matching the Hatch" (2006): "Matching the hatch ... can greatly increase your chances of success. That is not to suggest that it is every day on every river and lake that the trout feed in a selective way; sometimes all that the fish are looking for is food, ..."
 The 'selective-educated trout' makes a brief appearance under the subtitle "The hyper-cautious trout". "On heavily fished waters the largest trout may be battle-scarred survivors of many campaigns, continually on the look out for suspect flies. These hyper-cautious fish even reject a proportion of the natural flies that that drift past, perhaps because of a crumpled wing or a missing tail. And when large, succulent mayflies first appear they are often viewed with suspicion and shunned for quite some time. To catch an educated trout you will probably need a very close imitation of the natural creature it is feeding on at the time, ..." (O'Reilly 2006, p 18)
The 'selective-educated trout' makes a brief appearance under the subtitle "The hyper-cautious trout". "On heavily fished waters the largest trout may be battle-scarred survivors of many campaigns, continually on the look out for suspect flies. These hyper-cautious fish even reject a proportion of the natural flies that that drift past, perhaps because of a crumpled wing or a missing tail. And when large, succulent mayflies first appear they are often viewed with suspicion and shunned for quite some time. To catch an educated trout you will probably need a very close imitation of the natural creature it is feeding on at the time, ..." (O'Reilly 2006, p 18)
Other British writers remain wholeheartedly committed selectionists.
This quote is from British authors Peter Hayes and Don Stazicker in a Kindle book published in 2019: "Some writers have denied selectivity in trout, and criticised Swisher and Richards (“Selective Trout”, Crown Publishers 1971), notably Bob Wyatt (in “What Trout Want”, Headwater Books 2013). Our work puts us firmly among the believers in it." (Page 44) [emphasis added]
Bob Wyatt's book referred to by Hayes and Stazicker is a detailed critique of the terms 'the educated trout' and 'selectivity' as used by some British and American fly-fishing authors.
 The phrase 'selective feeding' is often portrayed as leading inexorably to 'educated trout' .
For example, Hayes and Stazicker (2019) provide this description of the relationship between learning / education, databases, selectivity and fly-tying:
The phrase 'selective feeding' is often portrayed as leading inexorably to 'educated trout' .
For example, Hayes and Stazicker (2019) provide this description of the relationship between learning / education, databases, selectivity and fly-tying:
"Given that we know that trout can learn from experiences ... it seems sensible to assume that every single successful and unsuccessful food capture all the way up from their being 1 inch long to the mature length at which we are trying to catch them, forms part of a database of experience that they use all the way through life."
"Now, that database of food items will, by the time we are trying to catch our trout in its maturity, amount to millions. And it is that database that we are trying to fit our imitation into in such a way that it matches the images of successfully eaten items, and does not jar." Hayes and Stazicker (2019 p. 63)
"... it is likely that trout can remember a savour [taste / smell] that may have accompanied last year’s BWO hatch, when it re-occurs this year." Hayes and Stazicker (2019 p. 66)
Maybe this hyperbole is just artistic licence. Trout can learn through experience (Johnsson & Kjällman-Eriksson 2008, Brown et al (2013), Reebs (2008), Kloepper 2016).
But viewing the trout's behaviour as involving a lookup table containing "every single successful and unsuccessful food capture...forms part of a database...they use all the way through life." ignores limits on storage, processing capacity, and the importance of forgetting as a crucial part of remembering (Schooler et al. 2012).
This can cause problems for humans as well as fish. In an uncertain environment, Too great a focus on information gathered from the past may retard effective decision-making about the future. Knowing too much can clog up the cognitive inbox, overload the neurological hard disk. (Haldane & Madouros 2012)
Hayes and Stazicker's (2019) approach leaves unanswered some fundamental questions, for example :
Failure to address this type of question fuels a false impression of the capabilities of an 'educated trout' 'inspecting' and 'rejecting' flies, which leads to this - quite unnecessary - need for precise imitation in artificial flies.
"The fly tyer needs to pay attention to the construction and materials of an imitation to get these matches right." (Hayes and Stazicker 2019, p63).
Interim summary
The prominence given to so-called 'selective or educated trout' can create the impression that selectivity becomes their modus operandi - a permanent habit ingrained by learned experiences.
Wyatt (2013) concludes that:
This may explain the apparent disconnect between current fly-fishing theory and contemporary behavioural sciences.
The popular view of 'PhD trout'
 "The growing numbers of skilled and well-equipped fishermen who crowd our rivers are educating a generation of PhDs. Trout are warier and harder to fool than ever." (Tapply 2017)
"The growing numbers of skilled and well-equipped fishermen who crowd our rivers are educating a generation of PhDs. Trout are warier and harder to fool than ever." (Tapply 2017)
These beliefs about the cognitive abilities of trout are not new. They were ridiculed 70 years ago by H.B. McCaskie in his book with the apt title 'The Guileless Trout' (1950 p 78) : "The belief, or delusion, that the trout is a highly intelligent creature is of comparatively modern origin, since it is a by-product of the development of the dry fly". McCaskie explains how followers of Halford found that when ever more precise imitations "..failed something had to be done in defence of injured pride, and the thwarted angler evolved the theory of a highly educated and shrewdly reasoning adversary ." McCasky realised that the trout's brain is incapable of these cognitive feats based on anthropomorphism. H.B. McCaskie's older brother, Norman, was a close friend of G.E.M. Skues so they were well-aware of Halfordian ultra-purist views.
Interim summary
The popular view of 'PhD trout' is a profound misconception on many levels that are explored in the rest of this article. Briefly, it implies that selective trout retain search images in long-term memory for use over their lifetime. In other words selectivity becomes the modus operandi for some trout. But this is very unlikely, a search image is a process employed to focus attention for a relatively short period of time whilst foraging for cryptic prey. To forage optimally predators need to be able to rapidly discard and replace search images to reflect changes in prey density (Ishii & Shimada 2009). Short-term memory is short-lived (usually from seconds to minutes) and labile; it can rapidly decay and can be easily erased by competing information.(Ishii & Shimada 2009).
In fact retaining 'selectivity' for a particular food item may be counter-productive: "Limited memory for food items may be related to the fact that in some environments, food sources can be quite variable, both in space and in time. There might be a trade-off (perhaps in terms of how many neurons can be allotted to each task) between remembering about old food sources, and learning to handle new ones. " (Reebs 2008)
The growing influence of ethology on fly design
It has been claimed that Halford transformed fly-fishing for trout into a scientific endeavour (Gubbins, 2018, Schweibert, 1979). I respectfully disagree. In the end, Halford laid out a rigid code for human conduct when fishing for trout. One element in Halford's dictum - precise imitation - was informed by entomology. But fly fishing subsequently developed "without the insight or understanding that the discipline of entomology could contribute to the subject." (Parrella 2013). Science has made an impact on fly-fishing equipment, but there has been a tendency in the last 50 years for some to decouple scientific insights into animal and fish behaviour, and even the accumulated wisdom of practitioners, from the endeavour. It's about time we faced up to these issues.
Many years ago Dr William Baigent (1862-1935) pointed out that advances in the scientific study of behaviour can provide important clues to the design of effective trout flies.
 In a letter to W. K. Rollo, Baigent wrote : "... the Baigent's Brown ...is based entirely on what the trout think themselves... It was made by finding out what combination of dry fly hackles would easily stimulate what Pavlov calls the trout's "investigating reflex", ... The fly will start such ocular reflexes when properly presented which will more often than not get the other reflexes in motion, and so the fly is taken in lamb-like fashion without suspicion and fuss." (Rollo, p 91-2, 1944 reprint).
In a letter to W. K. Rollo, Baigent wrote : "... the Baigent's Brown ...is based entirely on what the trout think themselves... It was made by finding out what combination of dry fly hackles would easily stimulate what Pavlov calls the trout's "investigating reflex", ... The fly will start such ocular reflexes when properly presented which will more often than not get the other reflexes in motion, and so the fly is taken in lamb-like fashion without suspicion and fuss." (Rollo, p 91-2, 1944 reprint).
Dr. Baigent is referring to Pavlov's [pictured] (1849-1936) award of the 1904 Nobel Prize in Physiology or Medicine for his studies of conditioned reflexes . We normally think of reflexes as singular events (e.g. knee-jerk reflex, pupillary light reflex, blink reflex etc). What is worth stressing is Baigent's suggestion that one reflex could trigger a chain reaction of reflex actions.
This may be the first mention of the the "chain reflex theory" of instinct in the fly-fishing literature. Baigent clearly had detailed knowledge of physiolgical studies of reflexes in the early 20th Century (Brigandt 2004), which may have influenced his attitude to Halfords dictum's about dry-fly fishing.
Baigent was in good company. One of the founders of modern ethology, and Nobel laureate Konrad Lorenz believed the chain reflex theory was a useful way to think about instinctive behavior, because the innate behavior patterns he studied were stereotyped which looks like a chain of reflexes. [Brigandt (2004) goes into the history of Lorenz's views on chain reflex theory.]
In his own way, Baigent anticipated what became called a Fixed Action Pattern (FAP) by ethologists (Lorenz 1981 p153). Reflexes still have a place in modern ethology. In his discussion of the difference between FAPs and reflexes the ethologist Burghardt (1973) commented: "it is likely that a continuum exists between isolated rigid reflexes (knee jerk, muscle twitch) and more complex and variable responses involving the whole organism."
 Bob Wyatt (2004 & 2013) pointed out that ethology - the study of animal behaviour - can make an important contribution to our understanding of trout feeding behaviour in terms of an adaptive evolved response, and showed how this can inform the design of effective trout flies.
Bob Wyatt (2004 & 2013) pointed out that ethology - the study of animal behaviour - can make an important contribution to our understanding of trout feeding behaviour in terms of an adaptive evolved response, and showed how this can inform the design of effective trout flies.
In his book The History of Fly Fishing in Fifty Flies Ian Whitelaw describes the thinking behind the Deer Hair Emerger: "For Wyatt, the goal of good fly design is not exact imitation but the presentation of one or more key triggers (shape, size and posture that form a general 'prey image') to elicit the feeding response...The Pheasant Tail Nymph and the Gold-Ribbed Hare's Ear clearly possess one or more of these key triggers .." (Whitelaw, 2015, p 207-8)

 Nikolaas Tinbergen ( 1907 – 1988) joined the Department of Zoology, University of Oxford in 1949. He developed ethological concepts - such as sign stimuli / 'triggers'/ key stimuli, search image, supernormal stimuli and fixed action patterns that today give us a scientific framework for understanding the success of fly-tying innovations such as the 'triggers' used by Wyatt, as well as older simpler patterns such as the Gold-Ribbed Hare's Ear and Frank Sawyer's Pheasant Tail Nymph.
Nikolaas Tinbergen ( 1907 – 1988) joined the Department of Zoology, University of Oxford in 1949. He developed ethological concepts - such as sign stimuli / 'triggers'/ key stimuli, search image, supernormal stimuli and fixed action patterns that today give us a scientific framework for understanding the success of fly-tying innovations such as the 'triggers' used by Wyatt, as well as older simpler patterns such as the Gold-Ribbed Hare's Ear and Frank Sawyer's Pheasant Tail Nymph.
In 1973 Tinbergen won the Nobel Prize for his discoveries. His work was made available to a wide audience through the book and film Signals for Survival made with fly-fishing author Hugh Falkus.
"Tinbergen found, it is often the case that quite crude tricks suffice, itself perhaps a reflection of animals’ greater reliance on simpler rules of thumb." i.e. heuristics (Hutchinson & Gigerenzer, 2005)
 The American writer of "A Modern Dry Fly Code", Vince Marinaro expressed it thus:"I am continually astonished by the fact that the most killing flies in fly-fishing history are of very simple construction" (Schullery, 1987 p232). This begs the question "Why did American fly-fishing authors remain wedded to using artificial flies that aim to be precise imitations of the natural insect especially when faced with large or mixed hatches?" The answer may lie in a difference in the attitude of
psychologists and zoologists to the roles of Nature and Nurture in the control of animal and human behaviours that permeated popular culture.
The American writer of "A Modern Dry Fly Code", Vince Marinaro expressed it thus:"I am continually astonished by the fact that the most killing flies in fly-fishing history are of very simple construction" (Schullery, 1987 p232). This begs the question "Why did American fly-fishing authors remain wedded to using artificial flies that aim to be precise imitations of the natural insect especially when faced with large or mixed hatches?" The answer may lie in a difference in the attitude of
psychologists and zoologists to the roles of Nature and Nurture in the control of animal and human behaviours that permeated popular culture.
Ethologists were zoologists, they focused on evolved, species-specific, innate / instinctive behaviours (i.e. Nature). They studied a wider range of animals - birds, fish and insects - usually under natural / field conditions.
 At that time research on animal learning in the UK, and the USA, was dominated by the American behaviourist Frank Skinner [pictured] (1904 –1990) . He is considered to be the most influential psychologist of the 20th century.
At that time research on animal learning in the UK, and the USA, was dominated by the American behaviourist Frank Skinner [pictured] (1904 –1990) . He is considered to be the most influential psychologist of the 20th century.
Behaviourists were psychologists who studied learning in rats and pigeons under controlled laboratory conditions and stressed the role of Nurture.
In the 1960s and early 70s ethologists and behaviourists clashed over their very different approaches to recording, analysing and interpreting behaviour. This tension is summed-up by the phrase Nature versus Nurture .
Behaviourists rejected the notion that instinct played a role in the development and expression of behaviours. Instead, they stressed the flexibility of behaviour shown by individuals, rather than the evolution of behaviour in species.
The aim of behaviourism was the discovery of general laws of learning that could be applied to all species including humans.
Behaviourists viewed the brain at birth as completely empty, a blank slate (tabula rasa). Learning / experience fills the void. There were no limits on what an animal could, or could not, learn about stimuli, responses and consequences. This so-called 'principle of equipotentiality' was at the heart of Skinnerian behaviourism but was later challenged in the 1970s.
In this video, Stanford University professor, Robert Sapolsky describes the sharp differences between the approach taken by ethologists and behaviourists to the study of animal behaviour.
He describes ethology as "interviewing an animal in its own language", an approach echoed in book titles by Datus Proper "What the Trout Said" and Bob Wyatt "What Trout Want".
 The prominence of behaviourism in the 1960- 70s may explain why best-selling fly-fishing authors such as Swisher and Richards in the USA, and Clark and Goddard in the UK reached the conclusion that 'educated' trout were difficult to catch because of a lifetime's learning experiences.
The prominence of behaviourism in the 1960- 70s may explain why best-selling fly-fishing authors such as Swisher and Richards in the USA, and Clark and Goddard in the UK reached the conclusion that 'educated' trout were difficult to catch because of a lifetime's learning experiences.
Influence of ethology on fly-fishing literature
 Before Wyatt's books(2004, 2013) and magazine articles (1999-2007) there was scarcely any mention of ethological concepts in the UK fly-fishing literature. Richard Walker briefly introduced the sign stimulus concept in his 1982 book Trout Fishing. It's surprising that there wasn't greater awareness in the UK given the prominence afforded by Tinbergen's Nobel Prize in 1973.
Before Wyatt's books(2004, 2013) and magazine articles (1999-2007) there was scarcely any mention of ethological concepts in the UK fly-fishing literature. Richard Walker briefly introduced the sign stimulus concept in his 1982 book Trout Fishing. It's surprising that there wasn't greater awareness in the UK given the prominence afforded by Tinbergen's Nobel Prize in 1973.
Curiously, Hugh Falkus - who worked with Tinbergen on their book and film Signals for Survival - did not refer to ethology when discussing the surface lure in his ground-breaking fly-fishing book Sea Trout Fishing published in 1978.
The situation was a little better in American literature.
 In 1994 Steeves and Koch's book Terrestrials devoted a chapter to ethological concepts - sign stimuli, fixed action patterns, learning, as well as search images. At the time Steeves was a Professor at Virginia Polytechnic Institute and State University. It probably helped that he was a biologist and removed from the academic debate between psychologists over Nature vs Nurture. But perhaps the implications of this chapter were not appreciated by a wider audience's preoccupation with mayflies.
In 1994 Steeves and Koch's book Terrestrials devoted a chapter to ethological concepts - sign stimuli, fixed action patterns, learning, as well as search images. At the time Steeves was a Professor at Virginia Polytechnic Institute and State University. It probably helped that he was a biologist and removed from the academic debate between psychologists over Nature vs Nurture. But perhaps the implications of this chapter were not appreciated by a wider audience's preoccupation with mayflies.
Furthermore, selectivity was less of a concern for the book's audience: "Luckily, the terrestrial fisherman encounters hatch selectivity only rarely" (Steeves and Koch, 1994 p 70)
But they "hit the nail on the head" with this comment: "It might be said that all the flies ever tied to imitate hatches have been formulated in the search for the correct sign stimulus for different situations" (Steeves and Koch, 1994 p 70).
Despite adoption of the word "trigger" to convey the sense of a sign stimulus, ethological analyses are still rare in the printed fly-fishing literature, but are increasingly found on the Internet (Rolston 2013, 2016 ). Skinnerian behaviourism remains a popular explanation for selectivity. For example, Hayes and Stazicker's conclusion (2019) [see above] that trout construct a ".. database of experience that they use all the way through life".
Are there limitations to behaviourism ?
Behaviourism's 'principle of equipotentiality' began to crumble in the 1970s. "The fact is that psychologists of learning have essentially ignored biological contributions to learning phenomena." (Schwartz, 1974)
In 1970 the prestigiuos journal Psychological Review published Martin Seligman's article On the generality of the laws of learning . It became a Citation Classic . Reflecting on its significance in 1980 Seligman wrote:
Video of the Garcia Effect: Conditioned Taste Aversion
"I intended it as an experimentely based attack on the tabula rasa principle. This was highly unpopular at the time, and Garcia, Paul Rozin, I, and others took considerable flak from traditional learning theorists. One major learning theorist said of Garcia’s findings, ‘They are no more likely to be true than you would find bird droppings in a cuckoo clock.’ The claim that natural selection might have influenced associability itself did not fall on wholly deaf ears, however. Younger learning theorists, ethologists, and cognitive psychologists under Chomskian influence found the claim congenial, if a bit ill-defined."
Seligman's theory of 'Preparedness of association' was a major theoretical breakthrough, it .. bridges a gap between the American learning tradition and the European ethological tradition, ...(Schwartz, 1974)
 In my experience, by the mid-late 1970s ethologists and behaviourists had became less divided by the Nature - Nurture debate. Both groups came to accept that genetic and environmental factors interact to control the development and expression of behaviour (Schwartz, 1974).
In my experience, by the mid-late 1970s ethologists and behaviourists had became less divided by the Nature - Nurture debate. Both groups came to accept that genetic and environmental factors interact to control the development and expression of behaviour (Schwartz, 1974).
The antecedents to essentially the same debate that erupted around ethology and sociobiology (behavioural ecology / evolutionary psychology) in the middle, and erupted again, at end of the 20th century are paraded - "warts-and-all" - by Barlow (1991); "At the heart of all this is the nature-nurture issue." (p294). And the debate about the mind as a 'blank slate' continues (Pinker 2004).
Is there a difference between 'educated' and 'selective' trout?
 The importance of evolution is now increasingly recognized in the American fly-fishing literature. But now evolution is presented as the explanation for selective educated trout. For example, "Evolution has made trout an extremely finicky, wary, and intelligent fish species, even though instinct, acute senses, and innate genetic programming are what trout possess, rather than humanlike intelligence." (Supinski 2014 p 61).
The importance of evolution is now increasingly recognized in the American fly-fishing literature. But now evolution is presented as the explanation for selective educated trout. For example, "Evolution has made trout an extremely finicky, wary, and intelligent fish species, even though instinct, acute senses, and innate genetic programming are what trout possess, rather than humanlike intelligence." (Supinski 2014 p 61).
In addition, if you "Combine these characteristics with catch-and-release and you get some of the most selective trout in the world" (Supinski 2014 p 61).
Supinski comments "Based on my personal experience, I find that for tough, selective trout on today's pressure waters, even the color of the thread and hook matter." (Supinski 2014 p 68).
I'm left wondering if selective trout are selective because of their evolutionary heritage, or because they learn to be selective in rivers that are popular with anglers.
The phrases "educated trout" , "selective trout" and "pressured trout streams" can pose difficulties. Wyatt (2013 p8-11) opines that "educated trout" and "selective trout" have reached the status of synonyms. The phrase "pressured trout streams" makes a brief appearance in Wyatt's discussion, but has a more prominent place in Supinski's (2014 p 68) explanation that experience of artificial flies causes selectivity on popular catch-and-release rivers.
It is perhaps not surprising that educated trout became a prominent feature of English chalk streams.
 In his last book Halford commented: "On a heavily fished water, trout are certainly more wary and more prone to come short than on a stream where the fisherman's presence is a comparative rarity." (Halford 1913 p150)
In his last book Halford commented: "On a heavily fished water, trout are certainly more wary and more prone to come short than on a stream where the fisherman's presence is a comparative rarity." (Halford 1913 p150)
I think Halford may have realised that this was, to some extent a problem of his own creation - the growth in popularity of dry-fly fishing: "..the spread of dry-fly fishing has become something dreadful to contemplate, because in the rivers where it is practised the fish never get a rest ..[and are] .. rendered more wary, more shy, and more suspicious.. " (Halford 1889 p42). He suggested that this led to successive generations of natural, and stocked, fish being less likely to feed on the surface.
He then went on to introduce an idea that was to become a central tenet of dry-fly fishing on English chalk streams: "The purists among dry-fly fishermen will not under any circumstances cast except over rising fish, and prefer to remain idle the entire day rather than attempt to persuade the wary inhabitants of the stream to rise at an artificial fly, unless they have previously seen a natural one taken in the same position." (Halford 1889 p42-3)
 But this idea was introduced cautiously because he immediately qualified his statement by adding: "Although respecting their scruples, this is, in my humble opinion, riding the hobby to death, and I for one am a strong advocate for floating a cocked fly over a likely place, even if no movement of a feeding fish has been seen there. " (Halford 1889 p43)
But this idea was introduced cautiously because he immediately qualified his statement by adding: "Although respecting their scruples, this is, in my humble opinion, riding the hobby to death, and I for one am a strong advocate for floating a cocked fly over a likely place, even if no movement of a feeding fish has been seen there. " (Halford 1889 p43)
In Halford's first book, I could find no reference to 'educated trout', 'wary', heavily fished, or trout coming short (via computer-based word search of Halford 1886). The word 'educated' to describe trout appeared in Halford's second book published in 1889, and was still in use a century later.
In 1985 I heard the word "educated" used to describe trout behaviour in a BBC TV programme with the title "The Educated Trout". In this segment Clarke and Goddard discuss using their new pattern - an Upside Down Paradun - to catch an educated trout. Subsequently, Clarke and Goddard used the phrase "experienced" rather than "educated" to describe "... fish that have seen plenty of artificial flies and learnt to avoid them." (Wyatt 2013, p 13) [emphasis added]. Clarke and Goddard developed a series of flies to deal with educated trout that I think, with the benefit of hindsight, cemented the unhelpful connection between "educated trout" and "selective trout".
I hope to show that "educated / experienced trout" and "selective trout" are not synonymous phrases. But first I need to address:
What are trout 'prepared' to learn ?
Learning ( / education / experience ) is important for an animal's survival. As well as structure, evolution has shaped what an animal can and cannot learn. It is now recognized that innate factors control learning especially when it has survival value (Garcia, 1955. Seligman & Hager, 1972. Schwartz, 1974. Pearce, 1997).
According to Seligman's (1970) theory of 'Preparedness' / 'Selective association' :
 A human example of 'preparedness' is the sauce bearnaise effect (a conditioned taste aversion). Seligman (1980) gives this description:
“Sauce bearnaise, an egg thickened tarragon-flavored sauce, used to be one of my favorite foods in the world. One evening, in 1966, I had sauce bearnaise on filet mignon. About six hours later I began to throw up and spent the next several hours retching. After that, sauce bearnaise tasted foul to me.“
A human example of 'preparedness' is the sauce bearnaise effect (a conditioned taste aversion). Seligman (1980) gives this description:
“Sauce bearnaise, an egg thickened tarragon-flavored sauce, used to be one of my favorite foods in the world. One evening, in 1966, I had sauce bearnaise on filet mignon. About six hours later I began to throw up and spent the next several hours retching. After that, sauce bearnaise tasted foul to me.“
Conditioned taste aversion has been found in fish (MacKay 1974), and birds (Lett 1980). It might be tempting to conclude that 'selectivity' in trout observed on popular rivers where anglers practice catch-and-release is an example of 'prepared learning'. Maybe these trout become selective as a result of learning that eating a fly is followed by being caught. For example: "As might be suspected, predatory fish spit out the prickly sticklebacks when they catch one. They also learn not to bother catching sticklebacks: after experience with sticklebacks, the pike predators' prey-catching sequence of eye movement, turn toward prey, stalk, leap, snap, turn prey head-forward in the mouth, and swallow, begins to be broken off at various points before the last two stages. " (Shettleworth 1974)
But the problem with invoking 'preparedness' as the explanation for a similar reaction by a trout to an artificial fly, is that a defining feature of prepared learning is 'selectivity'. Only a particular fly would be avoided in the future. Changing the fly should overcome the problem. Trout that have been caught and released continue to eat, and could be caught again. And, of course, only trout that have experienced catch-and-release would become selective.
A more plausible explanation for the difficulty catching trout on popular catch-and-release rivers is provided by considering the results of an experiment on learning by brook trout carried out by Mirza and Chivers (2000). Injured or disturbed fish release olfactory (smell) cues that serve as alarm signals, and convey to other fish the risk posed by a predator(s) in the vicinity. The alarm signal elicits a range of avoidance responses including fleeing and decreased foraging behaviour. The research showed that if a novel olfactory stimulus was presented together with an existing olfactory alarm signal, avoidance behaviour was elicited when the novel olfactory stimulus was presented on its own.
The effect:
Farnsley et al. (2018) have reported that pairing a visual cue (a potential predatory fish) with an olfactory alarm cue causes the visual cue to provoke avoidance behaviour in prey fish when presented on its own. This result supports the possibility that the olfactory cues from fish that are caught and released when paired with the visual cues of anglers might elicit avoidance behaviour(s) by trout.
 Research has shown that catch rates do decline on catch-and-release fisheries (Askey et al.2006; Young and Hayes 2004). Askey et al. suggest that "the population contained a group of highly catchable fish that were quickly caught and then learned to avoid hooks" (the same two fly patterns were used throughout the study ). The remaining fish in the population were deemed less vulnerable to angling effort. A similar distinction between more- and less vulnerable trout was made by Gubbins (2018) to explain the demise of dry-fly fishing for stocked trout in Michigan rivers. The costs could not be justified at the equivalent of an eye-watering $16-$62 per fish caught across several rivers. The problem arose because a lot of the stocked fish were not caught by anglers. Gubbins explains this in terms of the behaviour of stocked fish that were also deprecated by Halford writing in 1913: "When first thrown on their own resources they will take any fly offered to them...and a large proportion soon succumb to the wiles of the dry-fly fisherman.".
Research has shown that catch rates do decline on catch-and-release fisheries (Askey et al.2006; Young and Hayes 2004). Askey et al. suggest that "the population contained a group of highly catchable fish that were quickly caught and then learned to avoid hooks" (the same two fly patterns were used throughout the study ). The remaining fish in the population were deemed less vulnerable to angling effort. A similar distinction between more- and less vulnerable trout was made by Gubbins (2018) to explain the demise of dry-fly fishing for stocked trout in Michigan rivers. The costs could not be justified at the equivalent of an eye-watering $16-$62 per fish caught across several rivers. The problem arose because a lot of the stocked fish were not caught by anglers. Gubbins explains this in terms of the behaviour of stocked fish that were also deprecated by Halford writing in 1913: "When first thrown on their own resources they will take any fly offered to them...and a large proportion soon succumb to the wiles of the dry-fly fisherman.".
In a somewhat acerbic comment Gubbins (2018) wrote in Michigan "State fish managers schedule stocking operations for these rivers in May, thus offering the dry fly hero the opportunity in June to impersonate Halford on the Test thereby reliving the myth"
 Halford understood that stocking chalk streams is done for financial reasons.
Supinski (2014 p 3) relates how he and a friend caught 60+ trout on a stocked Test beat that resulted in the beat being closed for the rest of the season because the fish were uncatchable thereafter.
Halford understood that stocking chalk streams is done for financial reasons.
Supinski (2014 p 3) relates how he and a friend caught 60+ trout on a stocked Test beat that resulted in the beat being closed for the rest of the season because the fish were uncatchable thereafter.
On the two New Zealand rivers examined by Young and Hayes (2004) fish were either not seen after being caught and released or "were more likely to be scared by anglers or required smaller, low-profile flies before being caught than naïve trout. ".
I think learned avoidance of a predator (anglers) is a simpler explanation than that fish "learned to avoid hooks" (Askey et al.2006). It is revealing that both Supinski, O'Reilly and others (e.g. Deneki 2019) mention the need for stealth, concealment and careful presentation as ways of dealing with selective trout.
This is consistent with the suggestion that so-called 'selective' trout on popular venues have learnt to associate anglers with danger (Grand River 2015).
Of course trout can also learn to associate wading anglers with dislodged food, the so-called "San Juan Shuffle". Not all trout respond in this way to catch-and-release, Schill et al. (1986) reported that cutthroat trout were re-captured an average of 9.7 times during the 1981 season. But I've heard that they do have a reputation for being angler-friendly.
I detect growing acceptance of a clear distinction between "educated trout" and "selective trout". For example, "Educated trout are extremely good at picking up on the tell-tale signs of danger. When they’re alerted of danger, trout often will stop feeding and put up a guard." (Klewein 2020). Today the causes of, and ways of dealing with "educated trout" are very different from those advanced by Clarke and Goddard - tying flies to closely imitate natural flies. Nowadays, that approach is reserved for 'selective trout'.
Why do trout learn to be selective?
Background matching & prey detection
 On fast-flowing freestone Dartmoor rivers trout consume a wide variety of aquatic and terrestrial insects (Elliott, 1967).
In optimal foraging theory terms these trout are generalist predators.
On fast-flowing freestone Dartmoor rivers trout consume a wide variety of aquatic and terrestrial insects (Elliott, 1967).
In optimal foraging theory terms these trout are generalist predators.
Cryptic (camouflaged) prey are hard to detect. They are thought to have evolved coloured markings that match their background to avoid detection.
 Predators that hunt for cryptic prey learn "search images" for particular prey types (Dawkins 1971).
Predators that hunt for cryptic prey learn "search images" for particular prey types (Dawkins 1971).
Are trout similarly 'prepared' to enable them to learn to detect and consume insects with cryptic colours ?
Using the apparatus shown here, Johnsson & Kjällman-Eriksson (2008) investigated foraging by brown trout parr.
They comparing foraging:
Halford understood that stocking chalk streams is done for financial reasons.
Supinski (2014 p 3) relates how he and a friend caught 60+ trout on a stocked Test beat that resulted in the beat being closed for the rest of the season because the fish were uncatchable thereafter.Background matching & prey detection
On fast-flowing freestone Dartmoor rivers trout consume a wide variety of aquatic and terrestrial insects (Elliott, 1967).
In optimal foraging theory terms these trout are generalist predators.
Predators that hunt for cryptic prey learn "search images" for particular prey types (Dawkins 1971).
They comparing foraging:

 They found that:
They found that:
These results suggest that trout are generalist predators that form search images to survive in an environment with changing feeding opportunities such as diurnal variation in invertebrate drift.
The results are a good example of
the so-called 'evolutionary arms race' between predator and prey.
"Our results suggest that colour matching [ i.e. background matching ] may confer relatively long-term survival advantages [ for prey ] in natural systems as predator perceptive constraints seem to limit search image formation and / or search rate and, thus, foraging efficiency on the colour-cryptic prey." ( Johnsson & Kjällman-Eriksson, 2008) [emphasis added]
 The earliest and most famous study of 'background / colour matching' is
Kettlewell's experiment in the mid-1950s to study the evolutionary mechanism of industrial melanism in the peppered moth (Kettlewell 1959, Rudge 2005, Martinowksy 2007 larger version of image ).
The earliest and most famous study of 'background / colour matching' is
Kettlewell's experiment in the mid-1950s to study the evolutionary mechanism of industrial melanism in the peppered moth (Kettlewell 1959, Rudge 2005, Martinowksy 2007 larger version of image ).
"H. B. D. Kettlewell's work on the phenomenon of industrial melanism is widely regarded as the classic demonstration of natural selection and one of the most beautiful experiments in evolutionary biology." Not suprisingly it came under attack from .."advocates of creation and intelligent design".. (Rudge 2005).
 The results histogram shows the recapture rates of dark and light-coloured moths in heavily polluted and unpolluted forests.
The results histogram shows the recapture rates of dark and light-coloured moths in heavily polluted and unpolluted forests.
In summary:
It's worth making several points about these experiments involving birds (Kettlewell) and trout (Johnsson & Kjällman-Eriksson ) because they relate to our attempts to design effective trout flies.
The evolution of background matching in aquatic insects may protect them from predation ( Johnsson & Kjällman-Eriksson, 2008). If so, Halford's dictum of precise imitation of the colour, shape and size of natural insects is likely to result in artificial flies that are effectively camouflaged from a trout's point of view. This problem was identified by George La Branche in his 1914 book The Dry Fly and Fast Water.
'Background matching' may have implications for designing artificial flies. This comment by Swisher & Richards jumped out at me: "The right fly is one that resembles the natural so closely that the fish seem to prefer it to the real thing.. " [emphasis added]. Maybe the enduring success of some artificial flies is because they are more conspicuous - and therefore preferred to the natural fly they represent. The question remains: What can be added to the prey image to increase the conspicuousness of the artificial ?
Hunting by search image
 What is a 'search image? This example of human search images may help.
It was developed in the 1940s to train members of the The Royal Observer Corps Club to identify friendly and hostile aircraft. Recognition is based on a group of simple features - WEFT (Wingshape, Engine configuration, Fuselage shape and Tail type)
What is a 'search image? This example of human search images may help.
It was developed in the 1940s to train members of the The Royal Observer Corps Club to identify friendly and hostile aircraft. Recognition is based on a group of simple features - WEFT (Wingshape, Engine configuration, Fuselage shape and Tail type)
It is worth exploring the scientific origins of these terms, what features they may contain, how they enable animals (and humans) to pick out an object without checking it closely, as well as the current limits in our understanding. See (Hunting by Search Image ,Kenyon 2020).
 'Search image' was originally called "hunting by search image" by the ethologist Jacob von Uexkull in 1934. It enables an animal to concentrate attention on one prey item at a time.
'Search image' was originally called "hunting by search image" by the ethologist Jacob von Uexkull in 1934. It enables an animal to concentrate attention on one prey item at a time.
Predators develop a search image based on the prey that is encountered frequently. It can be replaced by a different search image as the density of prey changes and the animal shifts from one type of prey to another. Selective feeding in trout is probably the result of adopting a search image.
Most importantly, a search image is short-lived. It has a temporary existence. This distinguishes a search image from the persistent genetically-based response to a trigger / sign stimulus / sign stimuli (Ishii & Shimada 2009).
What is the difference between a 'sign stimulus', the plural 'sign stimuli, and a 'search image'? The differences are subtle . In trout species-typical behaviour (e.g. a 'rise') can be elicited by either sign stimuli, or a search image. In the absence of a search image, behaviour could be controlled by sign stimuli. There is an important and fundamental difference between these mechanisms.
By definition, in a newborn fish feeding behaviour is elicited by a sign stimulus, or a collection of sign stimuli. With maturation and practice each individual will form a search image. This search image will alter from time to time as an inevitable result of changing environment conditions, for example, the availability or absence of food.
What is a prey image?
'Let me introduce some ideas, just some things to kick around.' Quote from Gierach (1989).
 The term "prey image" (Aksnes and Giske 1993) was used by Bob Wyatt to discuss his ideas on fly design with friends (Dore (2009), Arden (2020) and Carl McNeil), and then present them to a wider audience in two books Trout Hunting (2004) and What Trout Want (2013).
The term "prey image" (Aksnes and Giske 1993) was used by Bob Wyatt to discuss his ideas on fly design with friends (Dore (2009), Arden (2020) and Carl McNeil), and then present them to a wider audience in two books Trout Hunting (2004) and What Trout Want (2013).
McDonnell (2016) adopted the (vulnerable) 'prey image' concept to design flies for salmon and pike as well as trout.
Wyatt and McDonnell maintain a clear distinction between the terms 'prey image' and 'search image':
Unfortunately, Wyatt's term prey image is sometimes misused as synonymous with search image (for example by Hayes and Don Stazicker 2019). All artificial flies are prey images. Their success depends on the extent to which they activate a trout's search image. We can only make an educated guess about what constitues a trout's search image and to how it varies from time to time.
Wyatt's Snowshoe Hair Emerger
 For Wyatt (2013 p 97) "What the trout responds to first are the most obvious aspects of the prey: size, shape and behaviour. If one of these essentials is missing or the fly behaves unnaturally, the lifeless artificial doesn't send the necessary "eat me " signals, the trout's go button isn't pushed, and it lets the fly go as "not food" ".
For Wyatt (2013 p 97) "What the trout responds to first are the most obvious aspects of the prey: size, shape and behaviour. If one of these essentials is missing or the fly behaves unnaturally, the lifeless artificial doesn't send the necessary "eat me " signals, the trout's go button isn't pushed, and it lets the fly go as "not food" ".
For Wyatt, as well as others, colour is a secondary consideration: "It is important only after the fly has passed the test as to size and profile." (Randall 2014).
Wyatt implies what Randall states explicitly: "A trout's search image has certain criteria our fly [i.e. prey image] must possess to be accepted." [emphasis added]
Randall adds: "The search image for floating insects and dry flies involves the footprint in the surface film and the correct sequence of visual clues as the insect or fly becomes visible in the window."
 If a trout fly - designed to be a prey image - contains all, or most of the, characteristics suggested by previous authors, that will cover all the bases, so to speak. Research suggests that a cluster of several factors increases the probability of a response (Figure 2 Section E in Bianco & Engert 2015). This is an example of the law of heterogeneous summation (Lorenz 1981 p184, Burghagen & Ewert 2017).
If a trout fly - designed to be a prey image - contains all, or most of the, characteristics suggested by previous authors, that will cover all the bases, so to speak. Research suggests that a cluster of several factors increases the probability of a response (Figure 2 Section E in Bianco & Engert 2015). This is an example of the law of heterogeneous summation (Lorenz 1981 p184, Burghagen & Ewert 2017).
In other words, the individual effects of several different 'triggers' in a prey image add up to increase the chances that a trout will rise to our fly.
The Recognition Heuristic
But I do wonder if a trout needs - or has the time in my local freestone rivers - to go through a checklist of properties (size, profile, colour, and behaviour) that must pass muster before accepting the artificial fly. That leads me to suggest that a quicker decision can be reached by using a simple "rule-of-thumb" - the recognition heuristic.
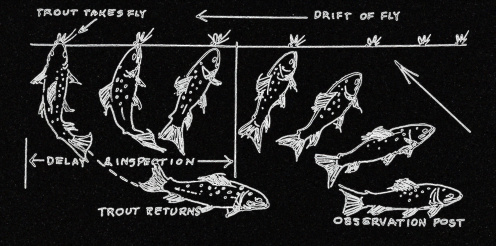 Why use a recognition heuristic rather than carrying out a slower but more accurate inspection of potential food? Using a recognition heuristic is more efficient in terms of optimal foraging theory (OFT).
Why use a recognition heuristic rather than carrying out a slower but more accurate inspection of potential food? Using a recognition heuristic is more efficient in terms of optimal foraging theory (OFT).
Handling time is an element in OFT which is a work-life balance model in behavioral ecology. Complex and compound rises are often given as examples of close inspection of natural and artificial flies. They may involve a tracking heuristic.
Recognition mistakes are not a threat to the animal's survival - trout don't die as a result of eating a particular insect. Mistakes can be rectified quickly - by spitting out the non-food item. (Neuswanger et al. 2014).
Pachur et al (2011) put it in this general way: "The key condition for the adaptive use of the recognition heuristic is its ecological rationality, when recognition accurately predicts the criterion in a given environment." The term "ecological rationality" refers to humans and other animals using simple strategies (rules-of-thumb / heuristics) to solve problems in complex environments (Todd and Gigerenzer 2012).
The recognition heuristic uses simple rules for searching, stopping the search, and decision-making. "Search is stopped whenever one object is recognized and the other is not; no further information is looked up about the recognized object." (Goldstein & Gigerenzer 2002). Therefore trout using a recognition heuristic do not need to engage in long 'inspections' of a fly in order to go through a checklist for the presence of every element in their search image before deciding to eat it.
 Put simply, lions don't count the stripes on both sides of the zebra.
Put simply, lions don't count the stripes on both sides of the zebra.
In a review of a decade of research on the recognition heueistic Pachur et al (2011) give this reason for my suggestion: "as long as the cue is correlated with other cues, ignoring this further knowledge minimizes the maximal deviation from the cue weighting scheme that could be derived under perfect knowledge of the environment. Importantly, this result does not depend on the one cue being the most valid one." [emphasis added]
Movement
 Movement (or the lack of it) is an example of trout responding to one element in a prey image, and ignoring others. During daylight trout will often shun unnatural drag on a fly. On dark nights brown trout and sea trout (Salmo trutta) will enthusiastically take a dragged fly, but refuse the same lure if it does not drag. Given the variety of materials - of different sizes and shapes - used to construct surface lures - once the wake of the dragged fly is recognized, presumably there is no further checking of size and shape etc. And it's very unlikely that some other factor in a prey image is capable of trumping unnatural drag in daylight.
Movement (or the lack of it) is an example of trout responding to one element in a prey image, and ignoring others. During daylight trout will often shun unnatural drag on a fly. On dark nights brown trout and sea trout (Salmo trutta) will enthusiastically take a dragged fly, but refuse the same lure if it does not drag. Given the variety of materials - of different sizes and shapes - used to construct surface lures - once the wake of the dragged fly is recognized, presumably there is no further checking of size and shape etc. And it's very unlikely that some other factor in a prey image is capable of trumping unnatural drag in daylight.
Colour
Colour - or more particularly the contrast between prey and its background i.e. conspicuousness (Aksnes and Giske 1993, Bianco & Engert 2015) - may be a useful factor when we construct prey images. Alternatively, colour may be an example of the opposite effect, but just as useful, an element in a prey image that trout may be willing to overlook, in favour of another element in the prey image (for example, size or shape).
My thoughts on the importance of colour are influenced by Johnsson & Kjällman-Eriksson's (2008) study of the effect of background matching & prey detection by brown trout parr, and Kettlewell's experiment on melanism in the peppered moth.
Johnsson & Kjällman-Eriksson found that brown maggots were easier to find by trout when presented against a green background, compared to brown maggots presented against a brown background. They varied the colour of the background, the colour of the prey (brown) was not altered. This leaves open the possibility that artificially-coloured green maggots may not have been eaten if they were presented against a brown background.
I suspect I'm just quibbling here about the need for a balanced design in this experiment. Fishing maggots are dyed a variety of colours, presumably to increase their conspicuousness.
Kettlewell's experiment avoids this problem. He used natural coloured moths (dark or light) presented against dark or light backgrounds. Kettlewell showed that birds captured more conspicuous moths than cryptic moths. Conspicuous insects were preferred over the cryptic version even though the birds were used to feeding on cryptic moths that matched the background. This shows that birds did not avoid novel coloured moths. In fact they preferred them presumably because they were easier to find. The birds ignored colour in their search image and instead used some other element(s) in their prey image (e.g. size and shape) My personal view is that precise colour matching may be less important than how conspicuous the prey image is against the background. There is a wide range of interesting opinions on the importance of colour in the fly-fishing literature.
A conspicuous prey image that triggers a recognition heuristic is a long way from Halford's dictum of a dry fly tied to be an exact imitation of an insect. But it seems to me to be an idea consistent with the ethologist Karl von Frisch's advice to Edward Hewitt in the early years of the last century.
I hope that thinking about trout flies as prey images that trout recognize as matching their search image will help overcome 'fly anxiety'. We all - from time to time - worry that we are not using the right fly. Tie or buy prey images rather than worrying about trying to slavishly match-the-hatch. Here is a collection of flies that have worked for me on local South Devon and Dartmoor rivers.
Designing prey images: Anti-Fly Tying & Supernormal / Superstimulus flies
"Anti-fly tying" is how Wyatt (2013 p 78) describes his approach to constructing prey images. Datus Proper (1993 p161) puts it more delicately, perhaps because he was about to confront a central tenet of fly-tying orthodoxy. Proper proposed a fundamental distinction between fly patterns and fly designs. Artificial flies were, and to a large extent continue to be, presented as 'patterns' consisting of a list of specific materials to tie onto a hook to represent wings, body, hackle etc. The reader is assumed to know where they go, and why they are there.
To illustrate the difference between pattern and design, Proper (p163) compares Halford with Baigent and Vince Marinaro:
 I am grateful to John Shaner for sending me a copy of Vince Marinaro's hand-drawn illustrations for tying his Thorax Dun, and a typed version of his hand-written instructions.
I am grateful to John Shaner for sending me a copy of Vince Marinaro's hand-drawn illustrations for tying his Thorax Dun, and a typed version of his hand-written instructions.
 Tom Mason's tying of Marinaro's Thorax Dun shows how it was designed to present a prey image consisting of:
Tom Mason's tying of Marinaro's Thorax Dun shows how it was designed to present a prey image consisting of:
In this YouTube video, Eric Austin ties Marinaro's Thorax Dun
But there are still influential dissenting voices.
Supernormal / Superstimulus trout flies
Marinaro addressed a fundamental problem associated with many artificial trout flies. Virtually all the trout flies in books, magazines, and on the internet will have at some time, under some circumstances, caught fish, but why they catch fish remains a mystery.
We have seen this uncertaintly expressed in different ways over the last century:
In my opinion, some flies contain sign stimuli that overcome the statistical problem identified by La Branche, and meet the desire expressed by Swisher and Richards for artificial flies that trout prefer to the real thing.
These are flies that have stood the test of time, and appear over-and-over again in lists of recommended flies. They may qualify for the ethological label 'supernormal stimulus' or 'superstimulus'.
"A supernormal stimulus or superstimulus is an exaggerated version of a stimulus to which there is an existing response tendency, or any stimulus that elicits a response more strongly than the stimulus for which it evolved." (From supernormal stimulus entry in Wikipedia ) [emphasis added]
I should use the word 'superstimulus' because my wife tells me 'supernormal' is an oxymoron !
 An important point that emerges from this YouTube video of a lecture by Deirdre Barrett, Ph.D. on "Supernormal Stimuli" is that humans may be good at creating superstimuli for other humans. But - unless we know the nature of the stimulus for which there is an existing response tendency - we cannot easily create superstimuli for animals. Consequently, very effective trout flies are more the product of serendipity than design. There's an obvious reason for this - we don't see the world from an animal's point of view (von Uexkull 1934). We tend to create flies that look like insects to us - that's the basis of Halford's precise-imitation dictum.
An important point that emerges from this YouTube video of a lecture by Deirdre Barrett, Ph.D. on "Supernormal Stimuli" is that humans may be good at creating superstimuli for other humans. But - unless we know the nature of the stimulus for which there is an existing response tendency - we cannot easily create superstimuli for animals. Consequently, very effective trout flies are more the product of serendipity than design. There's an obvious reason for this - we don't see the world from an animal's point of view (von Uexkull 1934). We tend to create flies that look like insects to us - that's the basis of Halford's precise-imitation dictum.
 This chapter in The Heuistic Trout describes how it is possible to create a 'supernormal stimulus' by modifying the size, colour and pattern of a sign stimulus.
This chapter in The Heuistic Trout describes how it is possible to create a 'supernormal stimulus' by modifying the size, colour and pattern of a sign stimulus.
The important message from these ethological experiments is that animals (and humans) prefer modified versions of natural objects, e.g. in terms of shape, size and colour (more specifically contrast - Burghardt 1973 p353).
 This allows fly tyers to add a 'soupçon' of 'je ne sais quoi' to a fly whilst relying on reassurance from the experts in selective attention that, just like the hook, if it isn't the superstimulus ingredient, it will be ignored because trout have a
"capacity for ignoring what it has no evolutionary reason to notice. " (Schullery 2006 p. 130).
At first, despite having an interest in ethology, I found it difficult getting used to flies that bore little or no resemblance to real insects. But experience has changed my mind.
This allows fly tyers to add a 'soupçon' of 'je ne sais quoi' to a fly whilst relying on reassurance from the experts in selective attention that, just like the hook, if it isn't the superstimulus ingredient, it will be ignored because trout have a
"capacity for ignoring what it has no evolutionary reason to notice. " (Schullery 2006 p. 130).
At first, despite having an interest in ethology, I found it difficult getting used to flies that bore little or no resemblance to real insects. But experience has changed my mind.
 Craig Matthews and John Juracek's (pictured) Sparkle Dun, is one of my
go-to flies for local (Devon UK) freestone rivers. In my opinion, it presents a prey image to trout. The Sparkle Dun contains the wing and thorax features thought to be responsible for triggering a trout's rise (Marinaro, Clarke & Goddard). In addition, it is relatively easy to tie, robust, and stands up to repeated encounters with trouts' teeth as well as repeated casting without the need for excessive false casting.
Craig Matthews and John Juracek's (pictured) Sparkle Dun, is one of my
go-to flies for local (Devon UK) freestone rivers. In my opinion, it presents a prey image to trout. The Sparkle Dun contains the wing and thorax features thought to be responsible for triggering a trout's rise (Marinaro, Clarke & Goddard). In addition, it is relatively easy to tie, robust, and stands up to repeated encounters with trouts' teeth as well as repeated casting without the need for excessive false casting.
 Craig Coltman's Antron Caddis, the Iris Caddis (pictured here), and the X-Caddis have similar properties. The Black X Caddis represents
a glossosoma caddis. The rivers running off Dartmoor are home to large numbers of this inconspicuous caddis. This page explores these lesser-known insects which can
be found on the top of stones in the rivers of Dartmoor and South
Devon.:
Some lesser-known West Country river flies
Craig Coltman's Antron Caddis, the Iris Caddis (pictured here), and the X-Caddis have similar properties. The Black X Caddis represents
a glossosoma caddis. The rivers running off Dartmoor are home to large numbers of this inconspicuous caddis. This page explores these lesser-known insects which can
be found on the top of stones in the rivers of Dartmoor and South
Devon.:
Some lesser-known West Country river flies
American flies are prominent in this collection of fly-tying videos that I find are effective on Dartmoor rivers. This is a 21st century example of the transatlantic sharing of fly-fishing ideas and techniques dating back to the 19th and 20th Century which is a theme of this essay. The Sparkle Dun was introduced to Devon anglers by Mike Weaver following his trip to Blue Ribbon Fishing in Southwest Montana.
These 'American-imports' are discussed in this informative video from John Juracek, Pat McCabe and Todd Tanner who have between them, " about 140 years of fly-fishing experience" - fishing for 'selective' trout on American rivers with world-famous fly hatches. They mention the need for a dry fly, and emerger, to be visible to the angler, and comment that a "chewed-up" fly is often more attractive to trout. They also mention "fishing the water" between hatches - a practice frowned upon by Halford in his later years - but the normal situation faced by anglers on UK freestone rivers. Pat McCabe freely admits that he enjoys tying flies that are pleasing to the eye. Many people feel this way, and it's perfectly understandable. Thankfully the days of strident Halfordian insistence on precise imitation, and social ostracism for transgressors, are disappearing in the rearview mirror of history. I recommend this long video because it reflects how Proper's distinction between pattern and design has been calmly absorbed into American fly-tying , but still leaves a place for a clear-eyed ascetically based choice of flies.
William G. Tapply sums it up this way: "We fly fishermen often marvel at the strange effectiveness of scraggly, bedraggled flies. An unused, beautifully tied imitation fresh from the fly box catches nothing until a wing falls off or the hackle starts unwinding. Suddenly trout cannot resist it. The reason is simple: Trout often key on scraggly, crumpled insects–cripples and stillborns and dead spinners–because they taste as good as the pretty ones and are easier to capture." Paul Schullery has some interesting thoughts on The Mystery of the Ratty Fly.

 Why are simple 'designed' flies effective, and some 'pattern' flies effective but complex? Maybe it's because a lot of pattern flies have the characteristics of a spandrel: twiddly bits added around sign stimuli / triggers.
Evolutionary biology borrowed the term spandrel from architecture to describe a structure or behaviour that is not particularly advantageous to have, but is retained because it is not particuularly harmful.
Why are simple 'designed' flies effective, and some 'pattern' flies effective but complex? Maybe it's because a lot of pattern flies have the characteristics of a spandrel: twiddly bits added around sign stimuli / triggers.
Evolutionary biology borrowed the term spandrel from architecture to describe a structure or behaviour that is not particularly advantageous to have, but is retained because it is not particuularly harmful.
Former owner of Blue Ribbon Flies, John Juracek (2020) writes in favour of flies being designed to solve a fishing problem, and offers an insider's explanation for the annual hatch of 'innovative' fly patterns. "There are still plenty of fishing problems awaiting solutions. Undoubtedly, some will be solved by better fly designs. But if our focus remains stuck on creating flies to solve business problems instead of fishing problems, it’s a safe bet that those solutions will be a long time coming."
Andrew Herd echoes Juracek's comment about new flies being produced to answer business influences, and adds: "In my opinion ninety-nine per cent of the new patterns we see are completely superfluous and they do fly fishing a disservice by confusing beginners and obscuring real innovation" (Herd 2003 p358).
Is the Gold-Ribbed Hare's Ear a Superstimulus ?
What was it that makes the Gold-Ribbed Hare's Ear(GRHE) such an effective artificial fly ?
It may be obvious, but the obvious is worth revisiting - the GRHE is the pinnacle of simplicity in artificial trout flies.
At some point Skues' wingless nymph became the dominant version of the GRHE. I think I'm safe in stating that nowadays the GRHE is sold and fished exclusively as a nymph. This is confirmed by the index in Hughes' weighty tome Trout Flies: The Tier's Reference [4lb 12 oz; 2.17Kg] which has no mention of the GRHE as a dry fly.
 Bob Wyatt uses hair from the hare's ear to create the body of his Deer Hair Sedge {pictured left} and Deer Hair Emerger [pictured right], flies which capture the essence of the Gold-Ribbed Hare's Ear - an effective dry fly abandoned by Halford, and adopted by Skues to represent a nymph.
Bob Wyatt uses hair from the hare's ear to create the body of his Deer Hair Sedge {pictured left} and Deer Hair Emerger [pictured right], flies which capture the essence of the Gold-Ribbed Hare's Ear - an effective dry fly abandoned by Halford, and adopted by Skues to represent a nymph.
The Deer Hair Emerger (DHE) and Deer Hair Sedge (DHS), are essentially modern versions of Halford's winged Gold-Ribbed Hare's Ear. The DHE is tied with a submerged abdomen to represent an emerger inspired by the Klinkhamer Special. Wyatt's designs back several horses; they represent cripples and emergers, as well as acknowledging the role of wing and thorax in triggering a rise (Marinaro, Clarke & Goddard) .
 For an ethologist, the spiky body in these three flies is a good candidate for a sign stimulus that enables trout to capture insects on, or trapped in, the surface film because :
For an ethologist, the spiky body in these three flies is a good candidate for a sign stimulus that enables trout to capture insects on, or trapped in, the surface film because :
Bob Wyatt's Deer Hair Emerger is essentially a modern winged version of Halford's abandoned fly - a winged Gold-Ribbed Hare's Ear tied as an emerger on a Klinkhamer hook.
Robert Smith traces the history of the use of hare's ear in artificial flies from Walton’s fifth edition of the Compleat Angler in 1676, through variations of a GRHE culminating in a Hare’s Ear Comparadun.
When I read Rob Smith's history several things stand out:
Is it possible that what a trout sees is hidden from us? Is there a property of fur from a hare's ear that only appears when it is wet? Ralph Cutter devotes a chapter in his book Fish Food (2005) to the range of insects that display gas bubbles. He makes an important about hare's ear fur, it can trap air bubbles. (Cutter p 43)
It's possible that the effectiveness of flies that have fur from a hare's ear as a body material is a result of its ability to trap air - a sign stimulus signalling food to trout.
The importance of presenting sign stimuli at an appropriate position in the water column
 As long ago as 1914 George La Branche put location within the water column in first position in his list of features that contributed to the success of an artificial fly. The primacy of 'position in the water' is still accepted today. Trout hunt for prey in particular locations. The next section provides a rationale (selective attention) for presenting a prey image in the location where trout expect to find a fly whether it be nymph, dun, emerger or spinner.
As long ago as 1914 George La Branche put location within the water column in first position in his list of features that contributed to the success of an artificial fly. The primacy of 'position in the water' is still accepted today. Trout hunt for prey in particular locations. The next section provides a rationale (selective attention) for presenting a prey image in the location where trout expect to find a fly whether it be nymph, dun, emerger or spinner.
How does 'selective attention' / search image control a trout's behaviour?
"We all see what we're looking for, and don't notice other things"
(Gingrich 1974 p304 quote from
" My Friend the Trout " by
Eugene V. Connett III)
Hinde (1970) points out that there is wide overlap between the concepts 'search image' and 'selective attention'. Subsequent research supports Hinde's attentional interpretation of the mechanism invoked by a search image (Bond and Kamil 1999).
 'Selective attention' refers to the process animals and humans use to filter information in their environment. Selective attention
promotes efficient decision-making in natural environments
by enabling animals to focus their attention on one, or a small number of stimuli, in order to accurately detect stimuli that, for example, signal food, and ignore irrelevant stimuli (Bater & Jordan 2019, Budaev et al 2019 ).
'Selective attention' refers to the process animals and humans use to filter information in their environment. Selective attention
promotes efficient decision-making in natural environments
by enabling animals to focus their attention on one, or a small number of stimuli, in order to accurately detect stimuli that, for example, signal food, and ignore irrelevant stimuli (Bater & Jordan 2019, Budaev et al 2019 ).
In humans it is called "The cocktail party effect" or'selective hearing'. It refers to the brain's ability to focus selective attention on a particular stimulus while filtering out a range of other stimuli. For example, focusing on a single conversation in a noisy room.
Incidentally, this explains why trout don't reject a fly tied on a hook. The hook does not signal food. The hook is ignored.
Attention has been interpreted as a process of stimulus selection produced by increases in the signal-to-noise ratio in specific sensory systems (Treisman, 1964 quoted in Brown and Warburton 1971).
I am proposing that trout use a search image to attend to a restricted number of sign stimuli that enables them to make quick decisions based on a prey recognition heuristic (rule-of-thumb) that saves their energy, and time (Andersen et al. 2016, Budaev et al 2019).
Hopefully this proposal will simplify your fly choice and fly design because:
trout do not look up information about previously eaten items in a database to decide whether or not to rise to your artificial fly (Hayes & Stazicker 2019)
it is not necessary for an artificial fly to be an exact imitation of a natural fly they have just eaten (Halford 1914, Swisher & Richards 2018, Supinski 2014)
Luuk Tinbergen (Niko Tinbergen's younger brother ) (1960) was the first to suggest that search images worked because of selective attention.
This generated a great deal of subsequent research that has been reviewed in detail by Kamil and Bond (2006). Kamil and Bond review laboratory studies that
"have firmly established the existence of the searching image effect" that develops after "multiple successive encounters with a single prey type" resulting in selective attention .
Here are three "take-home" messages :
I am proposing that trout use a search image to attend to a restricted number of sign stimuli that enables them to make quick decisions based on a prey recognition heuristic (rule-of-thumb) that saves their energy, and time (Andersen et al. 2016, Budaev et al 2019).
Hopefully this proposal will simplify your fly choice and fly design because:
"What is the relationship between sign stimuli and search images?"
I have an important unresolved question: "What is the relationship between sign stimuli and search images?" In my opinion, a search image involves a trout selectively attending to one, or more, sign stimuli to the exclusion of other sign stimuli.
Fish are born with a combination of sign stimuli that enables them to locate food items, and stereotyped movements that enable them to hunt and ingest prey.
Throughout its life, the characteristics of sign stimuli that a trout reacts to may alter as a result of maturation, and experience of food in its environment. For example, as a trout grows it needs to eat larger items of food. I discuss below experiments in which larval zebrafish approached small moving spots, larger spots elited turning-away avoidance behaviour. It is reasonable to suggest that the dimensions of spots that elicit approach and avoidance will increase with growth.
This change in responsiveness to sign stimuli may involve a mechanism that ethologists suggest filters responses to sign stimuli. The ethological concept of Innate Releasing Mechanism (IRM) enables an animal to respond to changes in available prey. The IRM is the gateway that is 'opened' by a specific sign stimulus, or set of sign stimuli, and 'releases' the Fixed Action Pattern (FAP) of behaviour that enables prey capture.
Burghardt (1973) reviewed the contribution of ethology to our understanding of instinctive behaviour. I consulted him for advice about the question at the start of this section. He pointed me towards this "rabbit-hole" to explore! " Wolfgang Schleidt published an important paper where he used not only IRM (Innate Releasing Mechanism) but also coined IRME (Innate Releasing Mechanism Modified by Experience). In fact, all sign stimuli can be modified to some extent by experience." (Gordon M. Burghardt, personal email 2020)
For example, visually guided prey-catching behaviour of the common toad - which is normally under the control of sign stimuli - can be modified by individual experience (Burghagen & Ewert 2017). A similar modification of sign stimuli was found in bitterling spawning behaviour (Heschl 1989)
At the bottom of this rabbit hole I found support for my suggestion that the IRM might provide a link between sign stimuli and search images. Unfortunately Schleidt (1962) was written in German, - beyond my ken.
But I found an English translation of a book written by Konrad Lorenz in which he describes Schleidt's suggestion that an IRM could be modified through experience to form an IRME. Lorenz adds this comment: "I must repeat that this process of increasing the selectivity of IRMs through learning is extremely common, not to say omnipresent. In fact, IRMs that are not adaptively modified by experience, thereby having achieved a higher degree of sensitivity, are not easy to find in higher vertebrates. " Lorenz (1981 p273-4)
Lorenz (1981 p184) makes two other pertinent points:
In addition, the stereotyped hunting behaviour of a young fish could be described as a fixed action pattern (FAP). But in an adult fish it is best described as a more fluid action - a Modal Action Pattern.
 I think we have reached a meeting point where concepts and insights from four academic disciples come together:
I think we have reached a meeting point where concepts and insights from four academic disciples come together:
Diagram from Burghagen & Ewert (2017)
 Reaction of the IRM and IRME to sign stimuli can be relied on to enable feeding in the absence of an abundance of a particular insect i.e. a trout feeding 'opportunistically' between hatches.
This set of sign stimuli can be likened to a box of chocolates. If any of the individual chocolates is encountered, it will be eaten. If an 'opportunistic' trout ate chocolates, they might pick a chocolate if it was either the appropriate size, shape, or colour. Our opportunistic trout is not a fussy chocolate eater.
Reaction of the IRM and IRME to sign stimuli can be relied on to enable feeding in the absence of an abundance of a particular insect i.e. a trout feeding 'opportunistically' between hatches.
This set of sign stimuli can be likened to a box of chocolates. If any of the individual chocolates is encountered, it will be eaten. If an 'opportunistic' trout ate chocolates, they might pick a chocolate if it was either the appropriate size, shape, or colour. Our opportunistic trout is not a fussy chocolate eater.
The chocolate box is my visual metaphor to illustrate Anne Treisman's first (1964) theory of selective attention: the attenuation theory of visual search. According to this theory, the signal-to-noise ratio for sign stimuli in the search image that are being attended to is increased, and therefore when encountered they stand out and elicit a trout's rise. In contrast, the signal-to-noise ratio for sign stimuli that do not form part of the search image is reduced, and those sign stimuli are ignored.
 A search image for a trout feeding 'selectively' involves the trout's IRME selectively attending to one sign stimulus configuration to the exclusion of other sign stimuli configurations.
That search image is like this picture of a box of chocolates with one chocolate being attended to. If this sign stimulus configuration (brown chocolate + heart shape) is seen it will be eaten.
A search image for a trout feeding 'selectively' involves the trout's IRME selectively attending to one sign stimulus configuration to the exclusion of other sign stimuli configurations.
That search image is like this picture of a box of chocolates with one chocolate being attended to. If this sign stimulus configuration (brown chocolate + heart shape) is seen it will be eaten.
 In the 1980's Treisman expanded her theory of selective attention: the feature integration theory of visual search (Treisman & Gelade 1980). It suggests there are two types of visual search: feature search and conjunction search.
In the 1980's Treisman expanded her theory of selective attention: the feature integration theory of visual search (Treisman & Gelade 1980). It suggests there are two types of visual search: feature search and conjunction search.
 My understanding of selective trout - as presented in the fly-fishing literature - is that they select prey on the basis of a conjunction search image that consists of several features that may include: size, shape, movement, colour "..the color of the thread and hook matter" (Supinski 2014)
My understanding of selective trout - as presented in the fly-fishing literature - is that they select prey on the basis of a conjunction search image that consists of several features that may include: size, shape, movement, colour "..the color of the thread and hook matter" (Supinski 2014)
This box of Quality Street chocolates illustrates the problem faced by a trout using conjunction search to select prey on the basis of size + shape + colour. It is a much slower process than feature search.
However, there is a solution that overcomes this problem, and speeds up the decision-making process. It's called the Recognition Heuristic.
 To introduce this solution, I've given our selective trout a penchant for Cadbury's heart-shaped chocolates. Now it's faced with a choice between 2 chocolates of the same colour, but different shapes - round, or heart shaped. In this situation, it will take longer for the trout to reach a decision because 2 features must be checked: colour plus shape, to ensure that it selects its preferred heart-shape chocolate.
To introduce this solution, I've given our selective trout a penchant for Cadbury's heart-shaped chocolates. Now it's faced with a choice between 2 chocolates of the same colour, but different shapes - round, or heart shaped. In this situation, it will take longer for the trout to reach a decision because 2 features must be checked: colour plus shape, to ensure that it selects its preferred heart-shape chocolate.
Faced with this situation, I would expect our chocolate-eating selective trout to use a recognition heuristic, and choose the chocolate on the basis of the first feature it recognises: colour or shape.
An example, based on a fly-fishing situation, may help to explain the Recognition Heuristic. When I'm guiding anglers, I advise them to use a fly they have confidence in, and after it lands on the water to "watch it like a hawk". This cuts through all the theoretical background. Hopefully it enables them to concentrate on simple things before a trout rises to their fly.
It's not always easy to see an artificial fly on the surface, especially if it's cast into the fast 'popply' water at the head of a pool. My advice is based on the probability that they already know a lot about the visual features of their familiar fly - its size, colour (contrast), where it should have landed (position / location), and movements (effect of current, or being taken by a fish).
 An experienced angler doesn't need to check all of these visual features one after another (size, location, movement and contrast) to spot their fly. They can rely on one visual feature (Treisman's feature search) to recognize their fly. A less experienced angler may need to use 2 or more visual features ( Treisman's conjunction search).
An experienced angler doesn't need to check all of these visual features one after another (size, location, movement and contrast) to spot their fly. They can rely on one visual feature (Treisman's feature search) to recognize their fly. A less experienced angler may need to use 2 or more visual features ( Treisman's conjunction search).
The trout's search image consists of one or more of the same set of simple visual cues to spot food: size, location, movement and contrast. Trout are very experienced in spotting potential food. They may be using a single feature as input to a recognition heuristic.
Archerfish shoot down insects using jets of water
In a laboratory, archerfish can be trained to shoot at target objects. Whenever the fish accurately shoots the target, they get a food pellet (Fazio 2020,Reichenthal et al 2020).

 Reichenthal et al (2020) found that archerfish performance on the search task was similar to human performance on the same task. Fish and humans found it easier to find the target if it had one feature that distinguished it from the background. For example, a red circle (the target) surrounded by blue circles of the same size. Time and accuracy did not increase as the number of distracting blue circles was increased. This is evidence of parallel visual search, Treisman's Feature search.
Reichenthal et al (2020) found that archerfish performance on the search task was similar to human performance on the same task. Fish and humans found it easier to find the target if it had one feature that distinguished it from the background. For example, a red circle (the target) surrounded by blue circles of the same size. Time and accuracy did not increase as the number of distracting blue circles was increased. This is evidence of parallel visual search, Treisman's Feature search.
 In contrast, fish and humans found the search task more difficult if the target consisted of two features that distinguished it from the background. For example, searching for a small blue circle from a background consisting of large blue circles, and small black circles. The time taken to locate the target increased as the number of distracting stinuli in the background increased. This is evidence of serial visual search, Treisman's Conjunction search
In contrast, fish and humans found the search task more difficult if the target consisted of two features that distinguished it from the background. For example, searching for a small blue circle from a background consisting of large blue circles, and small black circles. The time taken to locate the target increased as the number of distracting stinuli in the background increased. This is evidence of serial visual search, Treisman's Conjunction search
An angler who has confidence - based on experience with their fly - and a trout with a search image can both rely on the same recognition heuristic: a feature search image. The take-home message here is: you don't have to think like a fish, you are already thinking like a fish. You just haven't been told it yet.
Here is the link to an online demonstration of the difference in difficulty between feature and conjunctive searching.
This section in a previous essay suggests why selectively attending to one sign stimulus enables more efficient prey capture. A search image using a parallel (feature search) rather than a serial (conjunction search) visual process is an efficient and fast way of making a choice from items moving past trout in the drift.
"The parallel search mode occurs in the pre-attentive stage, where certain visual features are processed immediately throughout the entire visual field and in a hugely parallel fashion. If any of these features uniquely characterizes the target, the latter pops out to complete the search... a considerable amount of evidence [in humans and animals] indicates that features such as color, size, orientation, and motion make the target "pop out" of the display " (Reichenthal et al 2020).
Previous research (reviewed by Reichenthal et al 2020) has shown that
Reichenthal et al 2020 make this important point: "The literature on visual search in animals has (at least implicitly) supported the idea that anything that pops out for animals also pops out for humans, but not other way around. ". In other words things that "pop-out" for a trout may not "pop-out" for us. Jacob von Uexküll pointed out that each animal species lives within a sensory world created by the capabilities and limitations of their particular sensory systems (Burghagen & Ewert 2016).
In addition to using search images, trout may hunt for prey in particular locations, called 'hunting by expectation'. According to this theory animals learn to focus their foraging on an area that contains an abundance of food. This situation is familiar to fly-fishers. Insects are eaten at various stages in their life cycle from nymphs to spinners at different locations in the water column.
"If a forager has learned that the most common prey type is available in a specific microhabitat, then the cues associated with that habitat will provide a basis for associative priming. And, once it is hunting in the chosen microhabitat, the forager is likely to encounter the same prey type many times in succession, providing a basis for sequential priming" (Kamil & Bond 2006 p114).
In other words, trout may selectively search locations within the water column where they expect to find food.
What features are contained within a search image ?
It's all very well me saying that trout probably use a search image to detect prey, and that we stand a good chance of catching them if our artificial fly mimics that search image, but what feature(s) of the natural should we include and emphasize in our artificial flies?
It's important to recall that scientists, and fly-fishing authors, have remarked on the underlying simplicity of many animal behaviours. " Sign-stimuli [search images] are as simple as possible, and resemble the originals as closely as necessary." (Burghagen & Ewert 2017)
 But the search for simplicity is far from straightforward. Heschl (1989) describes the complexity involved in finding, and unravelling, innate (i.e. sign stimuli) and learned through experience components within the IRME (Innate Releasing Mechanism Modified by Experience) for mussel recognition in the European Bitterling. [ the bitterling spawns in the gills of a living freshwater mussel ]
But the search for simplicity is far from straightforward. Heschl (1989) describes the complexity involved in finding, and unravelling, innate (i.e. sign stimuli) and learned through experience components within the IRME (Innate Releasing Mechanism Modified by Experience) for mussel recognition in the European Bitterling. [ the bitterling spawns in the gills of a living freshwater mussel ]
We can only gain an uncertain into the sign stimuli that might be used in a trout's search image by 'reverse-engineering' from the features in effective prey images - effective trout flies. Suitable candidates include the Gold-Ribbed Hare's Ear and No-Hackle flies.
We just don't know for sure, but scientists who study visual search have given us clues that it might contain:
There is remarkable overlap between our current understanding of possible search image features, and the suggestions made by George La Branche in 1914 for features to incorporate in an artificial fly:
"My own experiences have convinced me that imitation of the natural insect is absolutely necessary, and I put the forms this should take in the following order — the order of their importance :
1st — Position of the fly upon the water, [i.e. location]
2nd — Its action [i.e.movement]
3rd — Size of the fly.
4th — Form [i.e. shape, appearance] of the fly.
5th — Colour of the fly."
These components are often found in contemporary descriptions of features to include in artificial flies. Swisher (2016) regards size and shape as more important than colour in artificial representatives of the natural fly. Hughes (1987) gives size, shape, colour, behaviour and habitat as critical factors in an artificial fly, and how it is fished.
Size and shape together with movement are listed in a standard textbook on animal behaviour.
"Most predators encounter a large number of different prey species that they have to discriminate from non-prey.
The three most commonly used cues are size, movement and shape." (McFarland 1985 p. 231)
Size and shape: Edward R. Hewitt (1866 - 1957)
 The importance of size and shape were recognised many years ago by the celebrated American author Edward R. Hewitt. He gave this piece of timeless advice: "I frequently change my fly for a similar one of another size and get good results, where other fishermen change the design of the fly itself. I have generally tried to change only one factor at a time, where possible. It would seem that I was acting on scientific principles in doing so. "(Hewitt 1948 p 54)
The importance of size and shape were recognised many years ago by the celebrated American author Edward R. Hewitt. He gave this piece of timeless advice: "I frequently change my fly for a similar one of another size and get good results, where other fishermen change the design of the fly itself. I have generally tried to change only one factor at a time, where possible. It would seem that I was acting on scientific principles in doing so. "(Hewitt 1948 p 54)
It's possible that Hewitt wrote this as early as 1922 because he refers to groundbreaking research by ethologist and Nobel laureate Karl von Frisch in the 1900s. At that time fish were thought to be color blind. von Frisch showed that fish were able to distinguish between objects based on their size, shape or colour, but importantly "They seemed to be able to take in one quality at a time, either color, shape, or size, but never two at a time. This corresponds with my own personal observation while fly fishing, where I have often found the size of the fly more determining of preference than design or color. " (Hewitt 1948 p 54)
Movement and vulnerability in a search image
I suspect that trout may develop a search image based on 'vulnerability'. Vulnerability is signalled by a lack of movement. A vulnerable insect is one that is unlikely to take off from the surface of the water for a variety of reasons identified by generations of fly fishers: an emerging dun, a dun trapped in its shuck, a spinner etc. The common feature here is 'lack of movement'. Sensitivity to movement is one of the features recognised from birth by larval zebrafish. This will present the trout with a vulnerable prey image.
How important is colour in a search image?
Before reading this section, I must prepare you with this perceptive comment from American fly-fishing historian Paul Schullery (1987, p229): "you must read a fair amount before you realize that you are hearing, in all these books [on fly-fishing theory], a disjointed and almost painfully polite debate".
There are examples of both ...
Opinions differ on the importance of colour. For example, Marinaro wrote this about imitating the colour of duns on the surface:
"The dun, riding lightly above the surface film, is never clearly defined at the point where the trout sees, inspects and takes the insect. Form and behavior are the most important elements. Any meticulous attention to color or detail is wasted effort” [emphasis added] (Marinaro, 1995 p26)."
 But Marinaro made a clear distinction about the importance of colour in representing flies lying on, or penetrating the surface:
But Marinaro made a clear distinction about the importance of colour in representing flies lying on, or penetrating the surface:
"Anything that breaks through the surface film is no longer obscured by the oblique rays or the diffusion above the [surface] film. Accordingly spinners, large terrestrials, emergers, and rising nymphs are extremely well defined as to color, form and parts. Special attention should be given to the remarkable transformation of the spinner wings as seen below the surface film..(op cit)
Dave Hughes remarked on the between-river and within-river variation in the colour of the same species of insect and commented: color varies "to such an extent that professional entomologists long ago threw out reference to colour as a legitimate way to accurately identify many of the insects." (Hughes 1987, p46)
Others go to great lengths to match the colour of the natural dun, and disagree with Marinaro.
For example, this from Hayes & Stazicker (2019) .
...we are in awe of his [Marinaro's ] groundbreaking work, but with the totally unfair benefit of 40-years-on digital full-colour underwater photography, we disagree: in the Plain Sight Zone of the window the fly is very clearly seen in colour as well as form, most of the time: frequently enough that we shall now always want to get the colour as right as we can...So the colours of flies are extremely important, since we want to maximise full congruence with prey image.”
(Hayes & Stazicker Page 311-312)
They quote this comment in “Dick Walker’s Trout Fishing”: “Conditions in which the trout cannot see the colours of the fly floating in his window must be rare indeed, if they ever exist.”
Hayes & Stazicker (Page 103) conclude:
"Both he [Dick Walker], and we, are sure that Marinaro’s conclusion that the flydresser can be content with imitating a) the footprint in the mirror and b) the wings, is wrong. Thorax hackle and tall wings are very far from sufficient to secure its commitment to eat the artificial. Our fly needs a body, and its body can and will be inspected closely, well lit, in the window, and therefore requires the flydresser’s full attention." (Hayes & Stazicker Page 103)
That's a lot of the accumulated knowledge in the American fly-fishing literature to throw out of the window! I'm reminded of Pascal's wager. I have examined Hayes & Stazicker's criticism of Marinaro's analysis of the role of the window and mirror in this section of a previous article.
Schullery offers this advice on what he calls 'fly theory': "Rule One: Don't insult the past. You may never realize the extent to which you are a product of it, ..."
My approach to colour in artificial flies is perhaps idiosyncratic:
- Cryptic colours have evolved to protect insects from predation. Therefore mimicking these colours of the natural insect in an artificial fly may be counterproductive.
- Kettlewell's experiment on colour / background matching showed that birds captured more conspicuous moths than cryptic moths. Conspicuous insects were preferred over the cryptic version.
-
Conspicuously coloured daphnids face a higher risk of predation from three‐spined sticklebacks (Raveh et al, 2018)
These results suggest that:
- Conspicuous colour did not disrupt bird and fish search images.
- Conspicuous colour could be incorporated in an artificial trout fly without disrupting the prey image.
These results suggest that:
- Conspicuous colour did not disrupt bird and fish search images.
- Conspicuous colour could be incorporated in an artificial trout fly without disrupting the prey image.
Of course, whilst colour may not be an essential part of a search image, there is no evidence that meticulously mimicking the colour of the natural insect will harm an artificial's compliance with the search image (i.e. Pascal's wager).
Will colour increase the conspicuousness of the artificial fly?
We are now in a better position to answer that question. Can we increase the conspicuousness of the prey image?
Why would we want to do this? For two reasons:
- La Branche pointed out that when an angler casts a traditional dry fly during a hatch of naturals - the statistical odds are stacked against the angler.
-
Swisher and Richards described the desirable feature in an artificial dry fly:
"The right fly is one that resembles the natural so closely that the fish seem to prefer it to the real thing" [emphasis added]. Swisher and Richards are describing what ethologists describe as a 'supernormal stimulus' - an artificial stimulus that are more effective than the real thing in eliciting a behavioural response. Would a conspicuous feature allied with a trigger / prey image meet their requirements?
 Unlike conventional trout flies, it's very difficult to design a supernormal artificial trout fly from scratch.
Conventional flies are designed with the natural fly as our model.
So-called 'attractor flies' may be the closest we have come to incorporating supernormal stimuli in a dry fly. It is interesting that some of these attractor flies have evolved at the hands of one or more fly tyers.
The results in the next section warns that a supernormal fly may look nothing like a natural.
Unlike conventional trout flies, it's very difficult to design a supernormal artificial trout fly from scratch.
Conventional flies are designed with the natural fly as our model.
So-called 'attractor flies' may be the closest we have come to incorporating supernormal stimuli in a dry fly. It is interesting that some of these attractor flies have evolved at the hands of one or more fly tyers.
The results in the next section warns that a supernormal fly may look nothing like a natural.
Could it be that the enduring success of some artificial flies is because they are prey images with an additional factor that makes them more conspicuous - and therefore preferred to the more subdued / cryptic / camouflaged natural fly?
 The Royal Wulff attractor pattern resembles no particular natural insect. Sometimes referred to as
a 'hatch-busting fly' to cope with selective feeding.
It is the result of developments made to a drab UK dry fly - the Coachman - that became popular in the USA. The modern version retains the double-hackle bi-visible introduced by E.R. Hewitt
. The hair-wing was added by the American Lee Wulff. An example of UK USA co-operation to produce a prey image with a conspicuous red band that may account for its long-lasting success.
The Royal Wulff attractor pattern resembles no particular natural insect. Sometimes referred to as
a 'hatch-busting fly' to cope with selective feeding.
It is the result of developments made to a drab UK dry fly - the Coachman - that became popular in the USA. The modern version retains the double-hackle bi-visible introduced by E.R. Hewitt
. The hair-wing was added by the American Lee Wulff. An example of UK USA co-operation to produce a prey image with a conspicuous red band that may account for its long-lasting success.
 Tup's Indispensable remains a popular and effective local (Devon) fly, with an international reputation. According to Whitelaw (2015), it inspired Skues' series of nymphs as well as American fly tyers: Leisenring, Hidy and Nemes. Precisely what natural fly it represents is unclear.
Tup's Indispensable remains a popular and effective local (Devon) fly, with an international reputation. According to Whitelaw (2015), it inspired Skues' series of nymphs as well as American fly tyers: Leisenring, Hidy and Nemes. Precisely what natural fly it represents is unclear.
The prescription for the mixture of colours (cream, orange, lemon, yellow, and crimson) and their proportions used to construct the thorax remained a commercial secret for several years after the inventor's death in 1911 (Courtney Williams 1979, Whitelaw, 2015, Kašpar, 2017).
 Maybe the colour of the thorax makes the artificial stand out from natural cryptic-coloured flies of a similar size and shape.
Local (Devonshire Avon) expert Cedric Potter even caught sea trout during the day on Tup's Indispensable. He fished it as a dry fly or nymph. It went through
several versions with increasing amounts of red / pink dubbing. I should explain. It's not just that sea trout are shy, fussy eaters. Sea trout generally don't eat anything on their return from the sea. They won't eat your, or anyone else's fly, even those created by the good Lord.
Maybe the colour of the thorax makes the artificial stand out from natural cryptic-coloured flies of a similar size and shape.
Local (Devonshire Avon) expert Cedric Potter even caught sea trout during the day on Tup's Indispensable. He fished it as a dry fly or nymph. It went through
several versions with increasing amounts of red / pink dubbing. I should explain. It's not just that sea trout are shy, fussy eaters. Sea trout generally don't eat anything on their return from the sea. They won't eat your, or anyone else's fly, even those created by the good Lord.
 Larvae of Ephemeroptera (mayflies), Odonata (dragonflies and damselflies), Plecoptera (stoneflies), Trichoptera (caddis), and Diptera ( chironomids, nonbiting midges, blood-worms) generally use red-coloured pigments (erythrocruorin or haemoglobin) to bind dissolved oxygen for respiration. (Hayashi 1989)
Larvae of Ephemeroptera (mayflies), Odonata (dragonflies and damselflies), Plecoptera (stoneflies), Trichoptera (caddis), and Diptera ( chironomids, nonbiting midges, blood-worms) generally use red-coloured pigments (erythrocruorin or haemoglobin) to bind dissolved oxygen for respiration. (Hayashi 1989)
Supernormal stimuli
 Supernormal stimuli is the phrase used to describe artificial stimuli that are more effective than the real thing in eliciting a behavioural response.
Supernormal stimuli is the phrase used to describe artificial stimuli that are more effective than the real thing in eliciting a behavioural response.
Herring gull chicks peck at a red spot on their parents's bill to induce their parents to regurgitate food. Chicks will also peck at a model consisting of a red spot against a yellow packground.
However it is possible to construct a model that is even more effective than a real head by using a red pencil with three white bars at the end. This is an example of a supernormal stimulus.
In this experiment the supernormal stimulus (the Stick) received about 25% more pecks from gull chicks than the natural head, a model of an adult head, or a model of the adult's bill (Tinbergen and Perdeck, 1950)
A description of an experiment demonstrating the law of heterogeneous summation is available on this page
Are trout born with the ability to hunt prey ?
Trout are probably not born with the ability to recognize and differentiate between chironomids, mayflies and stoneflies. This is not surprising given geographical variation in available food items across salmonid habitats and the size of juvenile fish. However, young salmonids do exhibit a set of probably innate feeding behaviours that enable them to approach objects in the drift, suck them in, spit them out, or ingest them (Neuswanger et al 2014). This section describes evidence that these behaviours are controlled by a set of sign stimuli. The interrelationship between sign stimulus, sign stimuli and search image is subtle, but important. Put simply a sign stimulus / sign stimuli control(s) the behaviour of all members of a species. The effects of search images vary across members of a species because they are formed as a result of an individual's experience.
The rest of this section describes recent research into identifying sign stimuli that control prey hunting in fish. It starts with an early classic experiment using toads. This is useful because it conveys how simple sign stimuli are, without involving baggage from the fly-fishing literature.
The scientific discipline neuroethology (Zupanc 2010) explores how sign stimuli are processed in the brain to control animal behaviour. The next video is based on an early classic study in neuroethology. The research involved several steps:

 This video
shows the visually guided prey-catching behaviour of the common toad. It demonstrates how the toad selects prey objects from non-prey on the basis of their size, movement and orientation (prey=horizontal versus non-prey=vertical). The main message here is the simplicity of the sign stimuli that release a complex behaviour (Ewert 1993, Burghagen & Ewert 2017).
This video
shows the visually guided prey-catching behaviour of the common toad. It demonstrates how the toad selects prey objects from non-prey on the basis of their size, movement and orientation (prey=horizontal versus non-prey=vertical). The main message here is the simplicity of the sign stimuli that release a complex behaviour (Ewert 1993, Burghagen & Ewert 2017).
Recent research shows that fish are born with an innate connection between sign stimuli and species-typical stereotyped behaviours that enable them to hunt and consume prey from very shortly after birth. The main aim of this research was to examine brain cell activity in real-time during prey capture. A first step was to select a stable behavioural baseline. The results of this element provide valuable insights into sign stimuli and stereotyped hunting behaviour.


When they start to swim zebrafish exhibit an innate ability to use simple sign stimuli to hunt and capture live prey (paramecium). After a successful capture, the larva assesses if it is food or not. If it is not judged to be food (e.g. an air bubble), the larva spits it out and swims away from it images here (Akira et al. 2013).
Bianco et al (2011) found that small moving spots evoke eye movements and J-turns of the tail, which are defining features of natural hunting in zebrafish at this age. Larger spots elited turning-away avoidance behaviour.
These effects have been replicated. Approach behaviour was elicited by small moving rectangles and avoidance behaviour when the size and velocity of the stimulus were increased. (Trivedi & Bollmann 2013) These results suggest that several sign stimuli movement, size and possibly shape control the behaviour.
The next question is "Are these features independent of each other? Is each feature capable of eliciting the hunting response on their own, or do they work in concert with each other?
Like previous research in toads, zebrafish hunting responses are triggered by a combination of visual features. "Size, contrast polarity [a bright spot on a dark background, or a dark spot on a bright background] and speed of motion interact, such that stimuli that are large, dark, and fast are most effective in triggering hunting responses." (Bianco & Engert 2015). The results that support this conclusion are available here
These results suggest that three sign stimuli control zebrafish hunting:
Thus several sign stimuli are more effective than a single sign stimulus in eliciting hunting behaviour in these fish.
The authors discovered 'feature-analyzing' brain cells that they suggest are responsible for detecting prey. These included neurons that responded to large, dark, moving spots - i.e. the most effective stimuli combination for eliciting hunting behaviour." We identified NLMS [Non-Linear Mixed Selectivity] neurons that preferentially respond to large, dark, fast-moving spots, which are the visual stimuli that were most effective in evoking hunting responses. These neurons are therefore candidates for mediating the perceptual recognition of optimal prey-like visual objects."
In addition, they found that dark spots had the greatest effect on eliciting hunting behaviour of any of the other single features they tested, and this was accompanied by finding "neurons that were highly selective for dark spots". Similar neurons in frogs and dragonflies have been described as "bug perceivers" by previous researchers.
Bianco & Engert (2015) suggest that because dragonflies and larval zebrafish attack prey from below "selectivity for dark targets might represent an adaptive feature of the visual system for discriminating prey against a relatively bright background." This is consistent with Goddard and Clarke's suggestion that the body of a dun in the trout's window is seen as a 'dark silhouette' rather than in colour. The importance of colour in trout flies is hotly debated.
In this clip from Ozzie Ozefovich's video Underwater World of Trout Part Two | Feeding Lies trout are feeding on the small mayfly Tricorythodes (Tricos)
The next question is "Do zebrafish continue to rely on a simple set of sign stimuli, or do they form a search image?". Predators develop a search image based on the prey that is encountered frequently. A search image will increase hunting efficiency. This has been demonstrated by Lagogiannis et al (2020). They studied the development of hunting behaviour in larval zebrafish by comparing hunting behavior (prey pursuit) and performance (prey capture) between larvae that had been reared with or without live prey.
They reported that:
The authors discuss the possibility that refinement of a heuristic 'rule-of-thumb' is responsible for development of improved hunting efficiency. " We cannot exclude the possibility that larvae posses an adaptive toolbox (see Todd and Gigerenzer, 2012; Todd and Gigerenzer, 2007) of distinct, preset, hunting strategies. This would imply that the role of learning is to utilize experience to find the best match between the set of available hunting behaviors and the particular foraging environments a larva encounters." Lagogiannis et al. (2020) [emphasis & link added]
Lagogiannis et al. (2020) are referring here to a startlingly simple way of explaining complex animal and human behaviours - heuristics [ Gigerenzer (2001);Hutchinson & Gigerenzer (2005); Gigerenzer & Gray (2017); Hamlin(2017) ] Heuristics enable "fast, frugal, and computationally cheap decisions. Heuristics are composed of building blocks that guide search, stop search, and make decisions." (Gigerenzer 2001)
In this video mathematics philosopher Gregory Wheeler explains how humans and animals use the Gaze - or Tracking - Heuristic to intercept moving objects without solving differential equations. And how the co-pilot Jeffrey Skiles used the Gaze Heuristic in the Hudson River incident
I have recently proposed that heuristics / rules-of-thumb are ideally suited for enabling prey capture by adult brown trout. (Kenyon 2020). I agree with Lagogiannis et al. (2020) that fish may also use a heuristic rule-of-thumb search image to recognize prey
Interim summary
What do mistakes tell us about search images?
Fly fishing depends on trout making mistakes, often and persistently !
 In freestone rivers :
"... all offerings must be quickly devoured and captured, since the trout's window of interception is small, and opportunity comes infrequently. It is interesting to stomach-pump a fish and investigate the ingestion process of high-gradient freestone stream trout. Often twigs, plastic/rubber bands and gravel might be found alongside aquatic invertebrates. The trout’s need to quickly intercept and capture food far outweighs the selective process." (Supinski 2014 p.10)
In freestone rivers :
"... all offerings must be quickly devoured and captured, since the trout's window of interception is small, and opportunity comes infrequently. It is interesting to stomach-pump a fish and investigate the ingestion process of high-gradient freestone stream trout. Often twigs, plastic/rubber bands and gravel might be found alongside aquatic invertebrates. The trout’s need to quickly intercept and capture food far outweighs the selective process." (Supinski 2014 p.10)
Heuristics may offer an explanation for this impetuous behaviour. Trout may be using a simple Recognition Heuristic to survive in their fast-moving environment. The Recognition Heuristic is part of an adaptive toolbox of distinct, preset, hunting strategies. The simple rule-of-thumb is to choose a recognized object. Searching is stopped whenever potential food is recognized. No further information is looked up about the recognized object (Goldstein and Gigerenzer 2002, Hutchinson and Gigerenzer 2005).
"Like the gaze heuristic, the recognition heuristic pays attention to only one variable—one which humans and animals have discovered through experience and have used to successfully deal with problems as varied as food choice and college choice. " (Gigerenzer and Wayne 2017)
I describe how trout may use a Recognition Heuristic to identify prey here in an earlier article.
 Trout need to react to potential food very quickly. Betts (2002) calculated that a trout lying 6 inches below the surface had one-eighth of a second or less, depending on the speed of the water, to react to food.
It is possible that trout recognize an edible object when they perceive an incomplete search image missing some element(s) size, shape, movement, or colour contrast.
Bianco & Engert's (2015) finding that hunting was elicited when a dark large stimulus was presented to larval zebrafish supports the suggestion that a sub-optimal set of sign stimuli is still capable of eliciting behaviour. Data available here. The brain cells responding to this feature were described as "bug percievers" in frogs and dragonflies.
This may account for 'mistakes' by trout. Furthermore trout have an innate mechanism for dealing with mistakes - their sense of taste.
Trout need to react to potential food very quickly. Betts (2002) calculated that a trout lying 6 inches below the surface had one-eighth of a second or less, depending on the speed of the water, to react to food.
It is possible that trout recognize an edible object when they perceive an incomplete search image missing some element(s) size, shape, movement, or colour contrast.
Bianco & Engert's (2015) finding that hunting was elicited when a dark large stimulus was presented to larval zebrafish supports the suggestion that a sub-optimal set of sign stimuli is still capable of eliciting behaviour. Data available here. The brain cells responding to this feature were described as "bug percievers" in frogs and dragonflies.
This may account for 'mistakes' by trout. Furthermore trout have an innate mechanism for dealing with mistakes - their sense of taste.
Young salmonids exhibit a set of innate behaviours: approach objects in the drift, suck them in, spit them out, or ingest them (Neuswanger et al 2014).
Neuswanger et al, (2014) used video cameras to record in 3D the feeding behaviour of juvenile Chinoook salmon during a four month period after they emerging as fry in an Alaskan river with potential trout food familiar to fly-fishers: chironomids, mayflies and stoneflies.
The fish displayed the ability to ingest, visually inspect and reject items in the drift. But the fish often made mistakes. They ingested a suprisingly large number of non-food items. The research revealed the importance of considering the impact of non-food debris is understanding salmonid feeding strategies.
Neuswanger et al (2014) reported: Among all potential food items fish pursued:
 The most interesting finding was the high percentage (52%) of items captured and quickly spat out after capture (see frequency chart), compared with the low percentage (9%) retained. Interestingly 39% of items were visually inspected and rejected.
The most interesting finding was the high percentage (52%) of items captured and quickly spat out after capture (see frequency chart), compared with the low percentage (9%) retained. Interestingly 39% of items were visually inspected and rejected.
This finding is confirmed by Jason Randall's (2014 p. 160) observation "During underwater filming, I have seen trout expel items faster than you can blink.
" ...The actual time the trout held some of the objects in their mouths was less than a second, which would make a hookset impossible even if the strike detection was immediate."
Similar 'mistake' behaviour has been seen in rainbow trout under laboratory conditions. " in the fine litter [natural substrate] treatment only about 71% of their attacks were directed toward prey; the rest were directed at pieces of litter about the same size and color as amphipods. " (Ware 1971a, p30) [emphasis added]. Ware also reported that adult rainbow trout fail to capture every prey they attack.
Randall offers a plausible explanation for 'mistake' behaviour: "the gustatory [taste] sense determines whether the trout will swallow the item or expel it. Even in visually oriented predators such as trout, a meal may be rejected based on taste once it is in the mouth.
... Younger fish with less experience will
experiment with objects by taking them into the mouth only to immediately expel them.
" Randall (2014 p. 160).
 All anglers miss fish. This is usualy put down to angler error; lack of attention or delay in striking. But it is possible that no matter how fast an angler strikes, the fish will have spat out the fly before any angler has time to react.
All anglers miss fish. This is usualy put down to angler error; lack of attention or delay in striking. But it is possible that no matter how fast an angler strikes, the fish will have spat out the fly before any angler has time to react.
The average reaction time for humans to a visual stimulus is 0.25 seconds. Given the time taken to raise the rod, take slack out of the line, and the speed of a trout's reaction time, it's not surprising that many fish are missed. The fly-fishing literature rarely includes estimates on the number of fish missed by anglers. Nicholas Fitton is a rare exception. He estimates that on freestone rivers in the North and West of the UK, rising 30 fish for every 10 caught is "pretty competent performance." Fitton (1992, p 144-6)
Even on chalk streams ".. a dry-fly fisher must expect to miss an abnormally large proportion of rises owing to the very small flies he uses .. " (Halford 1889 p45)
All anglers miss fish. This is usualy put down to angler error; lack of attention or delay in striking. But it is possible that no matter how fast an angler strikes, the fish will have spat out the fly before any angler has time to react.
An alternative explanation of 'inspection' and 'refusal'

In addition to a simple rise, Marinaro described two additional rise forms - compound and complex that were interpreted as evidence of the trout's close inspection of natural and artificial flies.
Marinaro makes the important point that compound and complex rises are seen in response to natural as well as artificial flies. And both types of fly can be refused. It's always surprised me that - having invested effort - a trout would refuse a natural fly. Is a simpler process involved? Recent neuroethological research may provide an answer.
Compound rise

"The compound rise is essentially a simple rise wherein the trout takes more time to examine the food source. It will continue to drift downstream with the fly and may even turn a little before it either takes or refuses it." Description from Fishing Small Flies By Ed Engle
Complex rise

"The complex rise occurs when the trout has considerable doubt about the food source. As the trout drifts downstream with the fly, it will stay further away from it and take longer to inspect it, it will either turn and quickly pursue the fly or refuse it. Vince Marinaro says that once the trout decides to turn downstream and chase the fly, he will never refuse it" Description from Fishing Small Flies By Ed Engle
In his description of the Compound rise Marinaro writes: "... in the compound rise the trout will stay with the insect (or artificial fly) throughout the entire drift, constantly inspecting while deciding whether to take or refuse." (Marinaro, 1995 edition, colour pages following text page 8)
Author of "Selectivity" Matt Supinski (p63) suggests that "Compound rises usually exist when the trout has been hooked before, has received substantial pressure on catch-amd-release waters, or there is a combination of phases in the hatch activity...".
I recently offered an alternative, albeit novel, explanation of a trout's apparent refusal after a period of inspection (Kenyon 2020).
During the downstream drift the trout's window is very small because the trout is close to the insect on the surface, "literally nose-to-nose with it" (Randall, undated). I suggested that breakdown in the trout's Tracking Heuristic may lead to so-called refusals. The trout's behaviour is similar to a sportsman using a 'modified gaze heuristic' to catch a ball (Todd and Gigerenzer 2012 p.7). For a trout this heuristic translates as: Fixate your gaze on the insect, start rising, and adjust your ascent so that the image of the insect approaches at a constant rate. If the insect approaches at a faster rate drop downstream to avoid the insect disappearing downstream out of your window . If the insect approaches more slowly, swim towards the prey.
Clip from "Floating Flies" (Nylander 2020)
Compound and complex rises are often reported on parts of a river where the surface is smooth, and the "fish have master's degrees in Selectivity" (Hughes 1987 p62). This behaviour is not restricted to limestone rivers or chalk streams. I have seen trout following a fly downstream in slow-moving pools on local freestone rivers.
But I have never seen a compound or complex rise on the faster flowing parts of these rivers where the water surface is uneven, and I doubt any one else has on similar water on free- or limestone rivers. The reason is simple, it would be very difficult to see compound and complex rises unless the surface of the water is relatively flat. But that does not exclude the possibility that trout in faster flowing parts of a river make compound and complex rises that we never see. Why would trout exhibit this behaviour?
Neuroethological research suggests that complex and compound rises are caused by trout adjusting their distance from prey prior to ingesting it, rather than 'inspecting' ot 'refusing' natural or artificial flies.
 Bianco et al's. (2011) findings on eye movements during hunting by larval zebrafish are consistent with this suggestion.
Bianco et al's. (2011) findings on eye movements during hunting by larval zebrafish are consistent with this suggestion.
 In a literature review Linton (2020) concluded "that vergence is one of our most important absolute distance cues for near distances. The consensus seems to be that “as targets get nearer, vergence information plays an increasingly important role in distance perception”, with vergence providing “critically important information” in reaching and grasping (Quinlan & Culham, 2007)."
In a literature review Linton (2020) concluded "that vergence is one of our most important absolute distance cues for near distances. The consensus seems to be that “as targets get nearer, vergence information plays an increasingly important role in distance perception”, with vergence providing “critically important information” in reaching and grasping (Quinlan & Culham, 2007)."
Diagram from Puig et al. (2010) Schematic explanation of the angle of eye vergence.
| The role of suction in ingesting a fly |
 Trout ingest insects of various sizes via suction which presumably requires very precise alignment - lining up with the fly to maximise the chance of sucking it into their mouth.
Trout ingest insects of various sizes via suction which presumably requires very precise alignment - lining up with the fly to maximise the chance of sucking it into their mouth.
The human equivalent of in-flight refueling, or threading a needle ... in the wind.
 Paul Schullery used this image in his book 'The Rise' to explain how
"The trout's suction feeding typically pulls a double-tapered column of
water into its mouth along with the prey".
Paul Schullery used this image in his book 'The Rise' to explain how
"The trout's suction feeding typically pulls a double-tapered column of
water into its mouth along with the prey".
The prey is the black dot in the "... center of the column at its greatest diameter..."
Is selectivity ubiquitous?
 I have never understood why the term 'selectivity' in the fly-fishing literature is reserved for 'educated' trout in slow-moving placid chalk streams during a large hatch of a particular insect. In freestone rain-fed rivers where food is less abundant trout are said to feed opportunistically.
I have never understood why the term 'selectivity' in the fly-fishing literature is reserved for 'educated' trout in slow-moving placid chalk streams during a large hatch of a particular insect. In freestone rain-fed rivers where food is less abundant trout are said to feed opportunistically.
I don't think it's helpful to draw any distinction between the mechanisms controlling the feeding behaviour of trout living in environments that vary in insect densities.
Are trout in freestone rivers selective feeders? Almost certainly. Why? Trout in freestone rain-fed rivers employ the basic search image
(a set of sign stimuli) to select edible items. In all rivers not everything in the drift is edible.
Conclusion: The future is full of possibilities
The sciences of (neuro)ethology and heuristics offer a way of understanding how trout select and catch their food, and potentially identifying the characteristics of effective trout flies that have stood the test of time.
Wyatt's (2013) criticism that
"For fly fishers, 'selective' and 'educated' trout are the same thing; the educated trout becomes more and more selective." (emphasis added) seems justified in light of the contrast between some fly-fishing literature, and what is known about the inherent simplicity of the mechanisms controlling fish feeding behaviours.
The ethologist Tinbergen pointed out that animal behaviours often turn out to be controlled by simple rules of thumb - heuristics. The 2002 Nobel Laureate Daniel Kahneman described heuristics as cognitive shortcuts or rules of thumb that simplify decisions, especially under conditions of uncertainty. There is a growing body of evidence that simple heuristics are used by human and animals (Gigerenzer and Gray 2017) and possibly trout to select and catch their prey (Kenyon, 2020).


About the author
Paul guiding ITV News reporter in June 2019
with sea trout in camera range ...
Paul Kenyon lives in Ivybridge on the southern edge of Dartmoor about 6 miles from the Upper Yealm Fishery.
Paul devotes more time than is reasonable to his love of all things associated with fish, fishing, instruction and guiding on Dartmoor rivers.
He retired in 2006 from the Department of Psychology, University of Plymouth where he lectured in behavioural neuroscience and evolutionary psychology.
email paul@flyfishingdevon.co.uk
Acknowledgement
In this video Bob Wyatt ties his Snowy Shoe Hare Emerger
Bob uses this material in place of CDC because he has found that CDC tends
to be "a one fish fly" which is an absolute no-no for guides on local rivers.
This article would not have been possible without the help and encouragement of Bob Wyatt.
Bob is an artist, author, Certified Fly Casting Instructor and long-time angler. Born in Canada, he fished the freestone streams of southwestern Alberta in the late 1950s. He now lives on New Zealand's South Island. His articles have appeared in Fly Fishing & Fly Tying(UK), Gray's Sporting Journal, Fly Rod & Reel, and Flylife Magazine (AU). He has published two books: Trout Hunting: The Pursuit of Happiness (2004) and What Trout Want: The Educated Trout and Other Myths (2013). In this interview by April Vokey he discusses his “prey image” theory, trout fishing and the early days of steelhead fly fishing.
Thanks to DASH: "A central, open-access repository of research by members of the Harvard community." for free open access to Bianco, I. H., Kampff, A. R., & Engert, F. (2011). Prey capture behavior evoked by simple visual stimuli in larval zebrafish. Frontiers in systems neuroscience, 5, 101.
References
-
Agro, Elisabeth R. (2000) "Caught by Trout, Piscatorial Books, and Fine Binding". The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 26, No 2, p 2-11, 2000. Available online
-
Akira Muto and Koichi Kawakami (2013). Prey capture in zebrafish larvae serves as a model to study cognitive functions. Neural Circuits, 11 June 2013. Available online
-
Aksnes, Dag L. and Jarl Giske,(1993). A theoretical model of aquatic visual feeding, Ecological Modelling, Volume 67, Issues 2–4, 1993, Pages 233-250,Available online
-
Andersen, Bjørn Snorre , Christian Jørgensen, Sigrunn Eliassen & Jarl Giske.(2016) "The proximate architecture for decision-making in fish". Fish and Fisheries. 2016,17, 680–695
-
Arden, Paul (2020). "The Good, The Bad and the Fugly". Sexyloops. Monday, 5 October 2020. Available online
-
Askey Paul J., Shanea Richards and John R Post (2006). "Linking Angling Catch Rates and Fish Learning under Catch-and-Release Regulations"
North American Journal of Fisheries Management26:1020–1029, 2006. Available online
-
Bater, Lovina & Jordan, Sara. (2019). Selective Attention. Available online
-
- Baerends , G. P. & Kruijt, J. P. (1973). Stimulus selection. - In: Constraints of learning (R. A. HINDE & J. STEVENSON-HINDE, eds), London (Academic Press).
-
Barlow George M. (1991). Nature-Nurture and the Debates Surrounding Ethology and Sociobiology, American Zoologist, Volume 31, Issue 2, April 1991, Pages 286–296, Available online
-
Belknap, Timothy. "George LaBranche's High Holt: A Place in His Life and Work" The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 18, No 2, 4, 1992.
Available online
- Betts, John. George La Branche: "A Very Beautiful Fisherman". The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 28, No 4, 12, 2002.
Available online
- Bianco, I. H., Kampff, A. R., & Engert, F. (2011). Prey capture behavior evoked by simple visual stimuli in larval zebrafish. Frontiers in systems neuroscience, 5, 101.
Available online
-
Bianco, Isaac H. & Florian Engert. (2015)
Visuomotor Transformations Underlying Hunting Behavior in Zebrafish,
Current Biology,
Volume 25, Issue 7,
2015,
Pages 831-846, Available online
-
Bond, Alan B. and Kamil, Alan C.(1999).Searching image in blue jays: Facilitation and interference in sequential priming. Animal Learning & Behavior.1999, 27 (4), 461-471.
-
Brigandt, Ingo (2004). "The Instinct Concept of the Early Konrad Lorenz". Available online
-
Brown, G.E., Ferrari, M.C.O., Malka, P.H. et al. Retention of acquired predator recognition among shy versus bold juvenile rainbow trout. Behav Ecol Sociobiol 67, 43–51 (2013).Available online
-
Brown, Kenneth and David Warburton (1971). "Attenuation ot stimulus sensitivity by scopolamine ". Psychon. Sci., 1971, Vol. 22 (5), 297-8.
-
Budaev Sergey, Jørgensen Christian, Mangel Marc, Eliassen Sigrunn, Giske Jarl
(2019)
"Decision-Making From the Animal Perspective: Bridging Ecology and Subjective Cognition"
Frontiers in Ecology and Evolution
VOLUME=7
YEAR=2019
PAGES=164 >Available online
-
Burghardt, Gordon. (2013). Effects of early experience on food preference in chicks. Psychonomic Science. 14. 7-8.
Available online
-
Burghardt, G. M. (1973). Instinct and innate behavior: Toward an ethological psychology. In J. A. Nevin & G. S. Reynolds, The study of behavior: Learning, motivation, emotion, and instinct. Scott, Foresman.
-
Burghagen, Harald & Ewert, Jörg-Peter. (2017). Burghagen H. and Ewert J.-P. (2016) Stimulus perception. Chapter 2 in: Bolhuis J.J. and Giraldeau L.-A. The Behavior of Animals: Mechanisms, Function and Evolution 2nd ed. Blackwell / Wiley. Submitted for publication. Available online
-
Clark, B. and J. Goddard (1980). The Trout and the Fly: A New Approach. London, Ernest Benn Ltd.
-
Clarke, Barry Ord. "The ultimate guide to deer hair part 2".
Available online
-
Clement, Skip (2019). "Is Schwiebert’s “Matching the Hatch” always best?" Fly Life, March 4, 2019. Available online
-
Clarke, Brian (1996). "Trout etcetera". A & C Black.
-
Cohen, Anna M. (2016). "Crazy Anglers: the rare breed of dry fly snobs" Blog entry April 20, 2016. Available online
-
Cooper, Simon & Charles Jardine (2020)."Hero vs. Villain. Halford or Skues". The Fishing Cast, Season 1 Episode 9.
Podcast available online
-
Courtney Williams, A Dictionary of Trout Flies, A & C Black. 1979
- Crowe, Sarah et al. The case study approach.
BMC Med Res Methodol. 2011; 11: 100. available online
-
Cutter, Ralph. Fish Food. Stackpole Books. 2005. Kindle Edition.
-
Dawkins, M. (1971). Perceptual changes in chicks: Another look at the 'search image' concept. Anim. Behav., 1971, 19, 566-574. Available online
-
DeLong, C. M., Keller, A. M., Wilcox, K. T., Fobe, I., & Keenan, S. A. (2018). Visual discrimination of geometric and complex 2D shapes in goldfish (Carassius auratus). Animal Behavior and Cognition, 5(3), 300-319.
Available online -
Deneki Outdoors Blog (2019). "5 Tips for Catching Educated Trout". Apr 29 2019. Available online
-
-
Dewar, George Albemarle Bertie, (1910). The book of the dry fly.
London : A. and C. Black Available online
-
Dore, Chris. (2009)"prey images and primary triggers". Blog available online
- Dukas Reuven, Alan C. Kamil, (2001) "Limited attention: the constraint underlying search image", Behavioral Ecology, Volume 12, Issue 2, March 2001, Pages 192–199, Abstract available online
-
Eckstein, Miguel P. ; Visual search: A retrospective. Journal of Vision 2011;11(5):14. doi: Available online
-
Elliott, J. M. (1967).“The Food of Trout (Salmo Trutta) in a Dartmoor Stream.” Journal of Applied Ecology, vol. 4, no. 1, 1967, pp. 59–71. JSTOR,Abstract available online Accessed 10 Apr. 2020.
-
Engle, Ed. Fishing Small Flies (2017). Stackpole .
-
Ewert, Jörg-Peter (1982). "Gestalt Perception in the Common Toad - 2. Modification of Prey Recognition by Learning".IWF \(Göttingen\. Available online
-
Ewert, Jörg-Peter {1993}. Image Processing in the Visual System of the Common Toad - Behaviour, Brain Function, Artificial Neuronal Net. IWF \(Göttingen\. Available online
-
Falkus, Hugh. (1978). "Sea Trout Fishing". H.F. and G. Witherby Ltd. (1978)
-
Farnsley Sarah, Kuhajda Bernard, George Anna, Klug Hope (2018). "Learning to Overcome a Lack of Evolutionary History: Can an Endangered Fish Learn to Fear an Introduced Predator?". Frontiers in Ecology and Evolution, vol 6 p 214 2018. Available online
-
Fazio, Lisa (2020). "Pop-out effects in visual search: Humans vs Archerfish". PsychonomicSociety. Available online
- Fitton, Nicholas. In Search of Wild Trout: Fly Fishing for Wild Trout in Rivers, Ward Lock, 1992.
-
Gaskell, Paul (2019) "Fly Fishing Master H.C. Cutcliffe Rediscovered in The Art of Trout Fishing in Rapid Streams". Published by the author.
-
Gerl, E.J., Morris, M.R. The Causes and Consequences of Color Vision. Evo Edu Outreach 1, 476–486 (2008)
-
Garcia, J.; Kimeldorf, D.J.; Koelling, R.A. (1955). "Conditioned aversion to saccharin resulting from exposure to gamma radiation". Science. 122 (3160): 157–158.
-
Gerl, E.J., Morris, M.R. (2008). The Causes and Consequences of Color Vision. Evo Edu Outreach 1, 476–486 (2008). Available online
-
Gierach, John (1989). "Fly Fishing Small Streams". National Book Network. Kindle edition. Limited content available online
-
Girard, Jerry (2003) "Thaddeus Norris: America's Izaak Walton"
The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 29, No 2, 3-7, 2003.
Available online
-
Goetz, Dave (2019).
"How Closely Should You Match the Hatch?". Published March 19, 2019. Blog article available online
-
Gierach, John. Fly Fishing Small Streams.Stackpole Books (1989)
- Gigerenzer, G. (2001). The adaptive toolbox. In G. Gigerenzer & R. Selten (Eds.), Bounded rationality: The adaptive toolbox (p. 37–50). The MIT Press. Abstract available online
-
Gigerenzer, Gerd and Daniel G. Goldstein (2011). "The recognition heuristic: A decade of research†" Judgment and Decision Making, Vol. 6, No. 1, January 2011, pp. 100–121
-
Gigerenzer, Gerd and Gray Wayne D. .(2017) Simple Heuristic Successfully Used by Humans, Animals, and Machines: The Story of the RAF and Luftwaffe, Hawks and Ducks, Dogs and Frisbees, Baseball Outfielders and Sidewinder Missiles—Oh My! Available online
-
Gigerenzer Gerd, Anja Dieckmann and Wolfgang Gaissmaier(2012),"Efficient Cognition Through Limited Search". Chapter 10 (p241-272) in Ecological Rationality: Intelligence in the World.
Peter M. Todd and Gerd Gigerenzer (Eds).
Print publication date: 2012
Print ISBN-13: 9780195315448.
Published to Oxford Scholarship Online: May 2012 Available online
-
Gingrich, Arnold. (1974) "The Fishing in Print: A guided tour through five centuries of angling literature". Winchester Press. Available online
- Goldstein Daniel G. and Gerd Gigerenzer (2002)
"Models of Ecological Rationality: The Recognition Heuristic". Psychological Review. 2002, Vol. 109, No. 1, 75–90. Available online
-
Gouws, Emeline Jeanne (2007)"Intraspecific body size frequency distributions" MSc thesis. Available online
-
Gouws EJ, Gaston KJ, Chown SL (2011) Intraspecific Body Size Frequency Distributions of Insects. PLoS ONE 6(3) Available online
-
Grand River Outfitting & Fly Shop (October 16, 2015). "Smart Trout?".Available online
- Gubbins, John (2018) "The Myth of the Dry Fly Hero: Its Rise and Fall in the Upper Peninsula of Michigan,"Upper Country: A Journal of the Lake Superior Region: Vol. 6 , Article 4.
Available online
-
Haldane, Andrew G. and Vasileios Madouros (2012)."The dog and the frisbee". Speech at at the Federal Reserve Bank of Kansas City’s 366th economic policy symposium, “The changing policy landscape”, Jackson Hole, Wyoming, 31 August 2012.Available online
- Halford, Frederic M. (1886)."Floating Flies and How to Dress Them". Available online
-
Halford, Frederic M. (1889). Dry-Fly Fishing in Theory and Practice. Available online
-
Halford, Frederic M. (1897). Dry Fly Entomology. London: Vinton & CoAvailable online
-
Halford, Frederic M. (1913). "The Dry-fly Man's Handbook: A Complete Manual, Including The Fisherman's Entomology and The Making and Management of a Fishery" . London: G. Routledge Ltd.
Available online
- Hamlin, Robert P.
“The Gaze Heuristic:” Biography of an Adaptively Rational Decision Process, Topics in Cognitive Science 9 (2017) 264–288
-
Hayashi, Fumio (1989) "Respiratory Responses of Aquatic Insects to Low Oxygen Concentration".
Japanese Journal of Limnology. Volume 50 (1989) Issue 3, 255-268.
Abstract available online
- Hayes, Peter and Don Stazicker (2019) Trout and Flies Getting Closer. Kindle Edition.
-
Herd, Andrew (2002). "Frederic M. Halford: The Myth and the Man". The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 28, No 1, 12-17, 2002.
Available online
-
Herd, Andrew Dr (2001). The Fly. Ellesmere, Shropshire: Medlar Press. pp. 304–5.
-
Herd, Andrew Dr (2003). The Fly. Ellesmere, Shropshire: Medlar Press.
-
Hewitt E.R. (1948). "A Trout and Salmon Fisherman for Seventy-Five Years". Charles Scribner's Sons.
-
Heschl, Adolf. (1989). Integration of “Innate” and “Learned” Components within the IRME for Mussel Recognition in the European Bitterling Rhodeus amarus (Bloch). Ethology 81, 193-208 (1989). Available online
- Hill J W (1930) "A Summer on the Test"
- Hinde, Robert (1970). "Animal Behaviour: A Synthesis of Ethology and Comparative Psychology". McGraw Hill.
-
Hubálek M. (2020). "A Brief (Hi)Story of Just-So Stories in Evolutionary Science". Philosophy of the Social Sciences. August 2020. Available online
-
Hughes, Dave (1987). "Handbook of Hatches: A basic guide to identifying local foods and selecting flies to match them". Stackpole Books.
-
Hughes, D. Trout Flies: The Tier's Reference. 1999. Stackpole Books.
-
Hughes, Dave (2005)."Handbook of Hatches: Introductory Guide to the Foods Trout Eat and the Most Effective Flies to Match Them". Stackpole Books.
-
Hutchinson, John M.C. , and Gigerenzer, Gerd. Simple heuristics and rules of thumb: Where psychologists and behavioural biologists might meet. Behavioural Processes 69 (2005) 97–124.
Available online
-
Ishii, Yumiko & Shimada, Masakazu. (2009). The effect of learning and search images on predator–prey interactions. Popul Ecol. 52. 27-35. Available online
-
Jacyna, Stephen 2009."The most important of all the organs:Darwin on the brain". Brain 2009: 132; 3481–3487. Available online
-
Johnsson, Jörgen & Kjällman-Eriksson, Karin. (2008). Cryptic prey colouration increases search time in brown trout (Salmo trutta): Effects of learning and body size. Behavioral Ecology and Sociobiology. 62. 1613-1620. 10.1007/s00265-008-0590-8. Available online
-
Juracek, John (2016).
"Does fly pattern matter? How often is fly choice your limiting factor".
Available online
-
Juracek, John (2020). "A Failure Of Modern Fly Design". Available online
-
Juracek, John (2020a). "A Sign Of The Times, Literally". Available online
-
Juracek John, Pat McCabe and Todd Tanner(2020b).In School of Trout. "Essential Dry Flies Video Class" Available online>
-
Kamil, Alan and Bond, Alan B., "Selective Attention, Priming, and Foraging Behavior" (2006). Papers in Behavior and Biological Sciences. Paper 40. Available online
-
Kašpar. Břetislav (2017) "Tup's Indispensable". Global Flyfisher. Published Mar 17. 2017. Available online
- Kenyon, Paul (2020) 'How does a trout catch a fly' Available online
-
Kenyon, Paul (May 2020) "How does a trout see a fly?". The Field, May 30th 2020.
Available online
-
Kettlewell H. B. D. (1959) "Darwin's Missing Evidence: Changes in Moths Observed" reprinted in Science on February 20, 2009. Available online
-
Kloepper (2016) Visual predators find cryptic prey more effectively by learning a search image. Available online
-
Klewein Kent (2020). "6 Easy Tips to Help Fly Anglers Catch Educated Trout". Gink & Gasoline Blog. Posted on June 16, 2020. Available online
- La Branche, George M L (1914) "The Dry Fly and Fast Water" Charles Scriber's Sons, New York. Available online at:
Available online
-
Lafontaine, Gary (1990)"The Dry Fly: New Angles". The Lyons Press.Sample available online
-
Lagogiannis K, Diana G, Meyer MP. (2020). Learning steers the ontogeny of an efficient hunting sequence in zebrafish larvae. Elife. 2020;9:e55119. Published 2020 Aug 10. doi:10.7554/eLife.55119 Available online
-
Lawton, Terry (2005). "Nymph Fishing a History of the Art and Practice". Stackpole Books. Elements available online
-
Lett, Bow Tong (1980). "Taste potentiates color-sickness associations in pigeons and quail". Animal Learning & Behavior 1980,8(2),193-198.
-
Linton. Paul (2020). "Does vision extract absolute distance from vergence?". Attention, Perception, & Psychophysics volume 82, pages 3176–3195(2020), Available online
-
Lorenz, Konard (1981)."The Foundations of Ethology", trans. Konrad Z. Lorenz and Robert Warren Kickert, New York: Springer Science+ Business. Preview available online
-
McCaskie, H.B. (1950). The Guileless Trout. Cresset Press.
-
McDonnell, Paddy. (2016) "The Vulnerable Prey Image (Part 2)". Blog entry March 19, 2016. Available online
-
McDonnell, Paddy. (2016) "The Vulnerable Prey Image (Part 1)". Blog entry March 19, 2016. Available online
-
McFarland David (1985).Animal Behaviour. Pitman.
-
MacKay, Barry (1974).
"Conditioned food aversion produced by toxicosis in atlantic cod",
Behavioral Biology,
Volume 12, Issue 3,
1974,
Pages 347-355.
Abstract available online
-
McNeil, Carl (undated). Interview in "Imitation or Presentation?". Midcurrent. Available online
-
Marinaro, V.C. (1970).A Modern Dry-Fly Code. Nick Lyons Books.
- Martinowksy (2007)
image of moths on tree. Attribution:
By Martinowksy, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=1855638
By Martinowksy, CC BY-SA 3.0
Link to larger version of image
-
Mason, Bill (1994). Fly Fishing: Learn from a Master. Sports Illustrated Books.
-
Mason, Bill (1994). Fly Fishing: Learn from a Master. Sports Illustrated Books.
-
Mirza, Reehan and Chivers, Douglas (2000). Predator-recognition training enhances survival of brook trout: Evidence from laboratory and fielden closure studies. Can. J. Zool.78: 2198–2208 (2000)
- Neuswanger Jason, Mark S. Wipfli, Amanda E. Rosenberger and Nicholas F. Hughes. (2014) Mechanisms of drift-feeding behavior in juvenile Chinook salmon and the role of inedible debris in a clear-water Alaskan stream. Environ Biol Fish (2014) 97:489–503 available online
-
Norris, T. (1865). The American angler's book: embracing the natural history of sporting fish, and the art of taking them. New edition. Philadelphia: E. H. Butler. Available online
-
Nylander, Rolf (2020). "Floating Flies | Full Film 2020". Available online
-
O'Reilly, Pat (2006). "Matching the Hatch: Stillwater, River & Stream". Swan Hill Press.
-
Overfield, (undated)."Skues and The World's Best Rod)". Flyfishers' Journal. p 96-8. Available online
- Pachur, Thorsten , Peter M. Todd, Gerd Gigerenzer, Lael J. Schooler and
Daniel G. Goldstein.(2011)
The recognition heuristic: a review of theory and tests Front. Psychol., 05 July 2011
-
Parrella, Michael P. (2013). "A History of the Entomology of Fly Fishing", American Entomologist, Volume 59, Issue 1, Spring 2013, Pages 15–27, Available online
-
Pearce, John M. (1997). Animal Learning and Cognition. Psychology Press.
-
Pilkington, David (2021) in Arundell Arms Hotel Spring 2021 newsletter,
-
Pinker, Steven (2004) "Why nature & nurture won’t go away". Dædalus Fall 2004: 5-17. Available online
-
Plaisted, K. C., & Mackintosh, N. J. (1995). Visual search for cryptic stimuli in pigeons: Implications for the search image and search rate hypotheses. Animal Behaviour, 50(5), 1219–1232.Available online
-
Proper, Datus.(1993) "What the Trout Said: About the Design of Trout Flies and Other Mysteries". Swan Hill Press.
-
Randall, Jason. "What Aboutt Refusals?", American Angler, undated.Available online
-
Randall, Jason. (2014) Trout Sense . Stackpole / Headwater. Kindle Edition. Stackpole Books.
-
Raveh, Shirley , Kathrin Langen, Theo C. M. Bakker, Christian Josephs, Joachim G. Frommen (2018). Oddity, predation risk and social decisions in aquatic invertebrates Ethology. 2018;1–8.
Available online
-
Reebs, Stéphan G. (2008) Long-term memory in fishes. Université de Moncton, Canada.Available online
-
Reichenthal, A., Segev, R. & Ben-Shahar, O. Feature integration theory in non-humans: Spotlight on the archerfish. Atten Percept Psychophys 82, 752–774 (2020).
Available online https://doi.org/10.3758/s13414-019-01884-4
- Rennie, James (1833) "Alphabet of Scientific Angling".
Available online
- Roberts, J (1994). To Rise a Trout.Marlborough, The Crowood Press.
-
Rollo, W. Keith (1944 ). Fly Fishing: Practical Hints on the Sport and Choice of Tackle and Water. H.F.& G.Witherby Ltd.,
-
Rolston, Tim (2016). "Vulnerability, A super stimulus?". The Fishing Gene blog. March 7th 2016 Available online
-
Rolston, Tim (2013) "The Cuckoo and the Trout". The Fishing Gene blog. August 31st 2013. Available online
-
Rolston, Tim (2020). "It’s NOT about the fly". The Fishing Gene blog. December 13th 2020. Available online
-
Ronalds, Alfred (1836)."The Fly-fisher's Entomology". Available online
-
Royal Entomological Society (2012). "Handbooks for the identification of British insects". Vol 1X Part 1.
Available online
-
Rudge, David W. (2005) The Beauty of Kettlewell's Classic Experimental Demonstration of Natural Selection. BioScience, Volume 55, Issue 4, April 2005, Pages 369–375.
Available online
-
Ruxton, Graeme D. , Tom N. Sherratt, and Michael P. Speed (2007)."Avoiding Attack: The Evolutionary Ecology of Crypsis, Warning Signals and Mimicry" Published to Oxford Scholarship Online: September 2007. Abstract & list of contents available online
-
Schill Daniel J. , J. S. Griffith & Robert E. Gresswell.(1986)" Hooking Mortality of Cutthroat Trout in a Catch-and-Release Segment of the Yellowstone River, Yellowstone National Park".North American Journal of Fisheries Management Volume 6, 1986 - Issue 2 Abstract available online
-
Schooler Lael J , Ralph Hertwig and Stefan M. Herzog (2012).
"How Smart Forgetting Helps Heuristic Inference".
Chapter 6 (p144-166) In
Ecological Rationality: Intelligence in the World.
Peter M. Todd and Gerd Gigerenzer (Eds).
Print publication date: 2012
Print ISBN-13: 9780195315448.
Published to Oxford Scholarship Online: May 2012 Available online
- Schullery, Paul (1987) "American Fly Fishing: A History" Lyons & Burford (p119-120).
-
Schullery, Paul (2002). "History and Mr. Gordon".
The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 28, No 1, 2-11, 2002.
Available online
-
Schullery, Paul (2007) "Norris on Trout Fishing: A Lifetime of Angling Insights". By Stackpole Books. Limited preview available online
- Schweibert, Ernest (1979) "Trout". Andre Deutch Ltd. London.
-
Schwartz, Barry. (1974). On going back to nature: A review of Seligman and Hager's Biological boundaries of learning. Journal of the Experimental Analysis of Behavior. 21. 10.1901/jeab.1974.21-183. Available online
- Seligman M E P. On the generality of the laws of learning.Psychol. Rev.77:406-18, 1970.
-
Seligman, M. E. P. & Hager, J. L. (1972, August). Biological boundaries of learning. The sauce-bearnaise syndrome. Psychology Today, V6, 59-61, 84-87.
- Seligman M E P. (1980). This Week’s Citation Classic. Current Contents Number 8, February 25, 1980. Available online
-
Shettleworth Sara J. (1974) .
Function, Causation, Evolution, And Development Of Behavior: A Review Of The Animal In Its World, By N.tinbergen.
Journal Of The Experimental Analysis Of Behavior.
1974, 22, 581-590.
Number 3 (November) Available online
-
Siebeck, Ulrike & Litherland, Lenore & Wallis, G. (2009). Shape learning and discrimination in reef fish. The Journal of experimental biology. 212. 2113-9. 10.1242/jeb.028936. Available online
-
Skues, G. E. M. (1921). The Way of a Trout with the Fly: And some further studies in minor tactics. London: Adams and Charles Black. Extract available online
- Skues, G. E. M. (1914). Minor Tactics of the Chalk Stream and Kindred Studies (2nd ed.). London: A & C Black. Available online
-
Smith, Robert (Undated). "Gold Ribbed Hare’s Ear". The Sliding Stream Blog. Available online
-
Steeves III, Harrison R. and Ed Koch. (1994). "Terrestrials: A Modern Approach to Fishing and Tying with Synthetic and Natural Materials". Stackpole Books.
-
Supinski, Matt (2014). Selectivity. Stackpole Books.
-
Sutcliffe, Tom (2013). "What fly rod did GEM Skues rate as his best and why?". Available online
"
-
Swentosky, Domenick (2019)."You don’t have to match the hatch". Troutbitten Blog. Jun 12, 2019. Available online
Swisher, D. & Carl Richards (1975). "Fly Fishing Strategies". Lyons & Burford.
-
Swisher, Doug (2016) Strategies for Selective Trout. YouTube video. Available here
- Swisher, Doug and Carl Richards (2018) "Selective Trout The Last Word on Stream Entomology and Aquatic Insect Imitation".
Kindle Edition
- Smith, Robert "Dr William Baigent".
Available online
-
Tapply, William G. (2016 reissue) “The Truth about Fly Fishermen,” Available online
- Tapply, William G. (2017 reissue)"How to Fool Really “Smart” Trout". Available online
-
The Fly fishers Club Spring Journal 1938 The Nymph Debate. Available on request
-
Todd PM, and Gigerenzer G. (2000). Précis of Simple heuristics that make us smart. Behav Brain Sci. 2000 Oct;23(5):727-41; discussion 742-80. Available online
-
Todd Peter M. and Gerd Gigerenzer (2012)."What Is Ecological Rationality?". Chapter 1 (p3-32) In
Ecological Rationality: Intelligence in the World.
Peter M. Todd and Gerd Gigerenzer (Eds).
Print publication date: 2012
Print ISBN-13: 9780195315448.
Published to Oxford Scholarship Online: May 2012 Available online
-
Travis, Tom (2016). "Skues, wet flies, nymphs and the angler".
Available online
-
Treisman Anne M. & Garry Gelade (1980)
"A feature-integration theory of attention". Cognitive Psychology, Volume 12, Issue 1, 1980, Pages 97-136. Abstract available online
-
Tinbergen, L. (1960). The natural control of insects in pinewoods. 1. Factors influencing the intensity of predation by songbirds. Archives Néerlandaises de Zoologie, 13, 265-343.
-
Trivedi,Chintan A. and Johann H. Bollmann (2013).Visually driven chaining of elementary swim patterns into a goal-directed motor sequence: a virtual reality study of zebrafish prey capture. Neural Circuits, 10 May 2013. Available online
-
von Uexkull, Jacob. (1934)"A stroll through the worlds of animals and men:A picture book of invisible worlds". Monograph available online
-
Ware, Daniel M. (1971a) The predatory behaviour of rainbow trout (Salmo gairdneri). PhD thesis. Department of Zoology, The University of British Columbia Vancouver 8. Canada
-
Weaver, Mike. (1991). "The Pursuit of Wild Trout". Merlin Unwin Books,
- Whitelaw, Ian (2015). 'The History of Fly Fishing in Fifty Flies' Arum Press.
-
Wikipedia (last edited 15 December 2020)."The Fly-fisher's Entomology". Available online
- Wyatt, Bob (2004). "Trout Hunting: The Pursuit of Happiness". Swan Hill Press.
- Wyatt, Bob. (2013). "What Trout Want: The Educated Trout and Other Myths" Stackpole Books.
-
Wyatt, Bob. (undated). "What Trout Want: A Beautiful Fiction". Excerpted from What Trout Want: The Educated Trout and other Fly-fishing Myths. Published in MidCurrant Available online
-
Young, Roger G. and Hayes, John W. (2004). "Angling Pressure and Trout Catchability: Behavioral Observations of Brown Trout in Two New Zealand Backcountry Rivers. North American Journal of Fisheries Management", 2004/11/01.
-
Zupanc, Günther K. H.(2010), Scholarpedia, 5(10):5306. Available online
Bibliography
- Bark, Conrad Voss (1992). "Halford". A History of Flyfishing. Shropshire, UK: Merlin Unwin Books. pp. 93–100. I
-
Barnard, Peter (2011) "Mayfly week: A brief history of fishing flies"
The Freshwater Blog. May 17, 2011. Available online
-
Birkhead, Tim.(2014)"Introduction to Animal Behaviour". YouTube video. Available online
- Burghardt, Gordon M. "Insights found in century-old writings on animal behaviour and some cautions for today". Animal Behaviour, Volume 164, 2020, Pages 241-249. Available online
-
Chomsky/Skinner Debate. YouTube video Available inline
- Dewar, George Albemarle Bertie, 'The book of the dry fly'
Available online
-
-
Dame Juliana Berners Collection. The American Museum Of Fly Fishing. Available online
- Gingrich, Arnold (1974). The Fishing In Print-A Guided Tour Through Five Centuries of Angling Literature. New York: Winchester Press.
-
Feature integration theory.Wikipedia. Available online
- Halford F M (1889) "Dry-Fly Fishing in Theory and Practice"
-
Fuchs, Jannon, Burghardt, Gordon. (1971). "Effects of early experience on the responses of garter snakes to food chemicals". Learning and Motivation. 2(3) 271-279. Available online
-
Goldstein, Daniel G. and Gerd Gigerenzer (2011). "The beauty of simple models: Themes in recognition heuristic research". Judgment and Decision Making, Vol. 6, No. 5, July 2011, pp. 392–395. Available online
-
Herbst, Ed (2012) "The Zakhåmer – Re-thinking the Klinkhåmer Special". Available online
-
Hewitt, Edward R. Hewitt's Handbook of Fly Fishing. New York: The Marchbanks press, 1933. Available online
-
McNamara JM, Houston AI. Integrating function and mechanism. Trends Ecol Evol. 2009 Dec;24(12):670-5. Abstract available online
-
Nastasi, Bob, and Al Caucci, (2004) Hatches II: A Complete Guide to Fishing the Hatches of North American Trout Streams.The Lyons Press.
-
Pachur, T., Todd, P., Gigerenzer, G., Schooler, L., & Goldstein, D. (2012). When is the recognition heuristic an adaptive tool.Available online
-
Pleskac, T. J. (2007). A signal detection analysis of therecognition heuristic.Psychonomic Bulletin & Review,14,379–391 Available online
-
Puig, M. & Zapata, L. & Moro, S. & Aznar-Casanova, J.Antonio & Supèr, Hans. (2010). Does eye vergence dissociate between covert and overt attention?. Journal of Vision - J VISION. 10. 157-157. 10.1167/10.7.157. Available online Source of Convergence (eyeballs) diagram
- Schullery, Paul (ed.) (2008) Skues on Trout: Observations from an Angler Naturalist. Mechanicsburg, Pa: Stackpole Books. pp. xi.
-
Shettleworth SJ. Taking the best for learning. Behavioural Processes. 2005 May;69(2):147-9; author reply 159-63. Abstract available online
-
Shettleworth SJ. Clever animals and killjoy explanations in comparative psychology. Trends in Cognitive Sciences. 2010 Nov;14(11):477-481. Abstract available online
-
Shaffer DM, Krauchunas SM, Eddy M, McBeath MK. How Dogs Navigate to Catch Frisbees. Psychological Science. 2004;15(7):437-441. Abstract available online
-
Schwiebert, Ernest. (2011) Matching the Hatch: A Practical Guide to Imitation of Insects Found on Eastern and Western Trout Waters. Revised edition 2011
- Seligman, M. E., & Hager, J. L. (1972). Biological boundaries of learning. Appleton-Century-Crofts.
-
Smith, Richard J. (2016) "Explanations for adaptations, just‐so stories, and limitations on evidence in evolutionary biology". Evolutionary Anthropology.
Volume25, Issue6, November/December 2016, Pages 276-287.
Abstract available online
-
Walker, Richard (1982). "Dick Walker's Trout Fishing" David and Charles.
-
Ware, Daniel M. (1971a) The predatory behaviour of rainbow trout (Salmo gairdneri). PhD thesis. Department of Zoology, The University of British Columbia Vancouver 8. Canada
-
Ware, Daniel M. (1971b). Predation by Rainbow Trout (Salmo gairdneri): the Effect of Experience.
Journal of the Fisheries Board of Canada
Volume 28, Number 12
1 December 1971.
Abstract available online
-
Wyatt, Bob. (2004). Dun and Dustered. Fly Fishing and Fly Tying. Available online
-
Wyatt, Bob. (2020). Fly Fishing Emergers. Available online
-
Wyatt, Bob. (c. 2003). Prime Cut. Fly Fishing and Fly Tying. Available online
-
Wyatt, Bob. (c. 2002-3). The Vulnerability Factor. Fly Fishing and Fly Tying. Available online
Attributions
John Juracek, Image of library book sale cart.
Patrón cocktail bar. Financial Times photo. licensed under the terms of the cc-by-2.0. Available online
Paramecium. By Barfooz at the English Wikipedia. - Originally uploaded to the English Wikipedia, where it was made by Barfooz., CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=172055
River Test.By Anthony de Sigley - Own work, Public Domain, https://commons.wikimedia.org/w/index.php?curid=34026185
River Test. By David Martin, CC BY-SA 2.0, https://commons.wikimedia.org/w/index.php?curid=73760844
San Antonio skyline. Bypassers, CC BY-SA 4.0 <https://creativecommons.org/licenses/by-sa/4.0>, via Wikimedia Commons"
Snowshoe Hair Emerger (SHE) image courtesy of Magnus Angus.
Spandrel figures of winged victories, Arc de Triomphe du Carrousel, Paris. By Thesupermat - Own work, CC BY-SA 3.0, Available online
The May Fly. Source: Extracted from public domain on the Internet Archive. Author Mike Cline.
About the author
Paul guiding ITV News reporter in June 2019
with sea trout in camera range ...
Paul Kenyon lives in Ivybridge on the southern edge of Dartmoor about 6 miles from the Upper Yealm Fishery.
Paul devotes more time than is reasonable to his love of all things associated with fish, fishing, instruction and guiding on Dartmoor rivers.
He retired in 2006 from the Department of Psychology, University of Plymouth where he lectured in behavioural neuroscience and evolutionary psychology.
email paul@flyfishingdevon.co.uk
Acknowledgement
In this video Bob Wyatt ties his Snowy Shoe Hare Emerger
Bob uses this material in place of CDC because he has found that CDC tends to be "a one fish fly" which is an absolute no-no for guides on local rivers.
This article would not have been possible without the help and encouragement of Bob Wyatt. Bob is an artist, author, Certified Fly Casting Instructor and long-time angler. Born in Canada, he fished the freestone streams of southwestern Alberta in the late 1950s. He now lives on New Zealand's South Island. His articles have appeared in Fly Fishing & Fly Tying(UK), Gray's Sporting Journal, Fly Rod & Reel, and Flylife Magazine (AU). He has published two books: Trout Hunting: The Pursuit of Happiness (2004) and What Trout Want: The Educated Trout and Other Myths (2013). In this interview by April Vokey he discusses his “prey image” theory, trout fishing and the early days of steelhead fly fishing.
Thanks to DASH: "A central, open-access repository of research by members of the Harvard community." for free open access to Bianco, I. H., Kampff, A. R., & Engert, F. (2011). Prey capture behavior evoked by simple visual stimuli in larval zebrafish. Frontiers in systems neuroscience, 5, 101.
References
-
Agro, Elisabeth R. (2000) "Caught by Trout, Piscatorial Books, and Fine Binding". The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 26, No 2, p 2-11, 2000. Available online
-
Akira Muto and Koichi Kawakami (2013). Prey capture in zebrafish larvae serves as a model to study cognitive functions. Neural Circuits, 11 June 2013. Available online
-
Aksnes, Dag L. and Jarl Giske,(1993). A theoretical model of aquatic visual feeding, Ecological Modelling, Volume 67, Issues 2–4, 1993, Pages 233-250,Available online
-
Andersen, Bjørn Snorre , Christian Jørgensen, Sigrunn Eliassen & Jarl Giske.(2016) "The proximate architecture for decision-making in fish". Fish and Fisheries. 2016,17, 680–695
-
Arden, Paul (2020). "The Good, The Bad and the Fugly". Sexyloops. Monday, 5 October 2020. Available online
-
Askey Paul J., Shanea Richards and John R Post (2006). "Linking Angling Catch Rates and Fish Learning under Catch-and-Release Regulations"
North American Journal of Fisheries Management26:1020–1029, 2006. Available online
-
Bater, Lovina & Jordan, Sara. (2019). Selective Attention. Available online
-
- Baerends , G. P. & Kruijt, J. P. (1973). Stimulus selection. - In: Constraints of learning (R. A. HINDE & J. STEVENSON-HINDE, eds), London (Academic Press).
-
Barlow George M. (1991). Nature-Nurture and the Debates Surrounding Ethology and Sociobiology, American Zoologist, Volume 31, Issue 2, April 1991, Pages 286–296, Available online
-
Belknap, Timothy. "George LaBranche's High Holt: A Place in His Life and Work" The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 18, No 2, 4, 1992.
Available online
- Betts, John. George La Branche: "A Very Beautiful Fisherman". The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 28, No 4, 12, 2002.
Available online
- Bianco, I. H., Kampff, A. R., & Engert, F. (2011). Prey capture behavior evoked by simple visual stimuli in larval zebrafish. Frontiers in systems neuroscience, 5, 101.
Available online
-
Bianco, Isaac H. & Florian Engert. (2015)
Visuomotor Transformations Underlying Hunting Behavior in Zebrafish,
Current Biology,
Volume 25, Issue 7,
2015,
Pages 831-846, Available online
-
Bond, Alan B. and Kamil, Alan C.(1999).Searching image in blue jays: Facilitation and interference in sequential priming. Animal Learning & Behavior.1999, 27 (4), 461-471.
-
Brigandt, Ingo (2004). "The Instinct Concept of the Early Konrad Lorenz". Available online
-
Brown, G.E., Ferrari, M.C.O., Malka, P.H. et al. Retention of acquired predator recognition among shy versus bold juvenile rainbow trout. Behav Ecol Sociobiol 67, 43–51 (2013).Available online
-
Brown, Kenneth and David Warburton (1971). "Attenuation ot stimulus sensitivity by scopolamine ". Psychon. Sci., 1971, Vol. 22 (5), 297-8.
-
Budaev Sergey, Jørgensen Christian, Mangel Marc, Eliassen Sigrunn, Giske Jarl
(2019)
"Decision-Making From the Animal Perspective: Bridging Ecology and Subjective Cognition"
Frontiers in Ecology and Evolution
VOLUME=7
YEAR=2019
PAGES=164 >Available online
-
Burghardt, Gordon. (2013). Effects of early experience on food preference in chicks. Psychonomic Science. 14. 7-8.
Available online
-
Burghardt, G. M. (1973). Instinct and innate behavior: Toward an ethological psychology. In J. A. Nevin & G. S. Reynolds, The study of behavior: Learning, motivation, emotion, and instinct. Scott, Foresman.
-
Burghagen, Harald & Ewert, Jörg-Peter. (2017). Burghagen H. and Ewert J.-P. (2016) Stimulus perception. Chapter 2 in: Bolhuis J.J. and Giraldeau L.-A. The Behavior of Animals: Mechanisms, Function and Evolution 2nd ed. Blackwell / Wiley. Submitted for publication. Available online
-
Clark, B. and J. Goddard (1980). The Trout and the Fly: A New Approach. London, Ernest Benn Ltd.
-
Clarke, Barry Ord. "The ultimate guide to deer hair part 2".
Available online
-
Clement, Skip (2019). "Is Schwiebert’s “Matching the Hatch” always best?" Fly Life, March 4, 2019. Available online
-
Clarke, Brian (1996). "Trout etcetera". A & C Black.
-
Cohen, Anna M. (2016). "Crazy Anglers: the rare breed of dry fly snobs" Blog entry April 20, 2016. Available online
-
Cooper, Simon & Charles Jardine (2020)."Hero vs. Villain. Halford or Skues". The Fishing Cast, Season 1 Episode 9.
Podcast available online
-
Courtney Williams, A Dictionary of Trout Flies, A & C Black. 1979
- Crowe, Sarah et al. The case study approach.
BMC Med Res Methodol. 2011; 11: 100. available online
-
Cutter, Ralph. Fish Food. Stackpole Books. 2005. Kindle Edition.
-
Dawkins, M. (1971). Perceptual changes in chicks: Another look at the 'search image' concept. Anim. Behav., 1971, 19, 566-574. Available online
-
DeLong, C. M., Keller, A. M., Wilcox, K. T., Fobe, I., & Keenan, S. A. (2018). Visual discrimination of geometric and complex 2D shapes in goldfish (Carassius auratus). Animal Behavior and Cognition, 5(3), 300-319.
Available online -
Deneki Outdoors Blog (2019). "5 Tips for Catching Educated Trout". Apr 29 2019. Available online
-
-
Dewar, George Albemarle Bertie, (1910). The book of the dry fly.
London : A. and C. Black Available online
-
Dore, Chris. (2009)"prey images and primary triggers". Blog available online
- Dukas Reuven, Alan C. Kamil, (2001) "Limited attention: the constraint underlying search image", Behavioral Ecology, Volume 12, Issue 2, March 2001, Pages 192–199, Abstract available online
-
Eckstein, Miguel P. ; Visual search: A retrospective. Journal of Vision 2011;11(5):14. doi: Available online
-
Elliott, J. M. (1967).“The Food of Trout (Salmo Trutta) in a Dartmoor Stream.” Journal of Applied Ecology, vol. 4, no. 1, 1967, pp. 59–71. JSTOR,Abstract available online Accessed 10 Apr. 2020.
-
Engle, Ed. Fishing Small Flies (2017). Stackpole .
-
Ewert, Jörg-Peter (1982). "Gestalt Perception in the Common Toad - 2. Modification of Prey Recognition by Learning".IWF \(Göttingen\. Available online
-
Ewert, Jörg-Peter {1993}. Image Processing in the Visual System of the Common Toad - Behaviour, Brain Function, Artificial Neuronal Net. IWF \(Göttingen\. Available online
-
Falkus, Hugh. (1978). "Sea Trout Fishing". H.F. and G. Witherby Ltd. (1978)
-
Farnsley Sarah, Kuhajda Bernard, George Anna, Klug Hope (2018). "Learning to Overcome a Lack of Evolutionary History: Can an Endangered Fish Learn to Fear an Introduced Predator?". Frontiers in Ecology and Evolution, vol 6 p 214 2018. Available online
-
Fazio, Lisa (2020). "Pop-out effects in visual search: Humans vs Archerfish". PsychonomicSociety. Available online
- Fitton, Nicholas. In Search of Wild Trout: Fly Fishing for Wild Trout in Rivers, Ward Lock, 1992.
-
Gaskell, Paul (2019) "Fly Fishing Master H.C. Cutcliffe Rediscovered in The Art of Trout Fishing in Rapid Streams". Published by the author.
-
Gerl, E.J., Morris, M.R. The Causes and Consequences of Color Vision. Evo Edu Outreach 1, 476–486 (2008)
-
Garcia, J.; Kimeldorf, D.J.; Koelling, R.A. (1955). "Conditioned aversion to saccharin resulting from exposure to gamma radiation". Science. 122 (3160): 157–158.
-
Gerl, E.J., Morris, M.R. (2008). The Causes and Consequences of Color Vision. Evo Edu Outreach 1, 476–486 (2008). Available online
-
Gierach, John (1989). "Fly Fishing Small Streams". National Book Network. Kindle edition. Limited content available online
-
Girard, Jerry (2003) "Thaddeus Norris: America's Izaak Walton"
The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 29, No 2, 3-7, 2003.
Available online
-
Goetz, Dave (2019).
"How Closely Should You Match the Hatch?". Published March 19, 2019. Blog article available online
-
Gierach, John. Fly Fishing Small Streams.Stackpole Books (1989)
- Gigerenzer, G. (2001). The adaptive toolbox. In G. Gigerenzer & R. Selten (Eds.), Bounded rationality: The adaptive toolbox (p. 37–50). The MIT Press. Abstract available online
-
Gigerenzer, Gerd and Daniel G. Goldstein (2011). "The recognition heuristic: A decade of research†" Judgment and Decision Making, Vol. 6, No. 1, January 2011, pp. 100–121
-
Gigerenzer, Gerd and Gray Wayne D. .(2017) Simple Heuristic Successfully Used by Humans, Animals, and Machines: The Story of the RAF and Luftwaffe, Hawks and Ducks, Dogs and Frisbees, Baseball Outfielders and Sidewinder Missiles—Oh My! Available online
-
Gigerenzer Gerd, Anja Dieckmann and Wolfgang Gaissmaier(2012),"Efficient Cognition Through Limited Search". Chapter 10 (p241-272) in Ecological Rationality: Intelligence in the World.
Peter M. Todd and Gerd Gigerenzer (Eds).
Print publication date: 2012
Print ISBN-13: 9780195315448.
Published to Oxford Scholarship Online: May 2012 Available online
-
Gingrich, Arnold. (1974) "The Fishing in Print: A guided tour through five centuries of angling literature". Winchester Press. Available online
- Goldstein Daniel G. and Gerd Gigerenzer (2002)
"Models of Ecological Rationality: The Recognition Heuristic". Psychological Review. 2002, Vol. 109, No. 1, 75–90. Available online
-
Gouws, Emeline Jeanne (2007)"Intraspecific body size frequency distributions" MSc thesis. Available online
-
Gouws EJ, Gaston KJ, Chown SL (2011) Intraspecific Body Size Frequency Distributions of Insects. PLoS ONE 6(3) Available online
-
Grand River Outfitting & Fly Shop (October 16, 2015). "Smart Trout?".Available online
- Gubbins, John (2018) "The Myth of the Dry Fly Hero: Its Rise and Fall in the Upper Peninsula of Michigan,"Upper Country: A Journal of the Lake Superior Region: Vol. 6 , Article 4.
Available online
-
Haldane, Andrew G. and Vasileios Madouros (2012)."The dog and the frisbee". Speech at at the Federal Reserve Bank of Kansas City’s 366th economic policy symposium, “The changing policy landscape”, Jackson Hole, Wyoming, 31 August 2012.Available online
- Halford, Frederic M. (1886)."Floating Flies and How to Dress Them". Available online
-
Halford, Frederic M. (1889). Dry-Fly Fishing in Theory and Practice. Available online
-
Halford, Frederic M. (1897). Dry Fly Entomology. London: Vinton & CoAvailable online
-
Halford, Frederic M. (1913). "The Dry-fly Man's Handbook: A Complete Manual, Including The Fisherman's Entomology and The Making and Management of a Fishery" . London: G. Routledge Ltd.
Available online
- Hamlin, Robert P.
“The Gaze Heuristic:” Biography of an Adaptively Rational Decision Process, Topics in Cognitive Science 9 (2017) 264–288
-
Hayashi, Fumio (1989) "Respiratory Responses of Aquatic Insects to Low Oxygen Concentration".
Japanese Journal of Limnology. Volume 50 (1989) Issue 3, 255-268.
Abstract available online
- Hayes, Peter and Don Stazicker (2019) Trout and Flies Getting Closer. Kindle Edition.
-
Herd, Andrew (2002). "Frederic M. Halford: The Myth and the Man". The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 28, No 1, 12-17, 2002.
Available online
-
Herd, Andrew Dr (2001). The Fly. Ellesmere, Shropshire: Medlar Press. pp. 304–5.
-
Herd, Andrew Dr (2003). The Fly. Ellesmere, Shropshire: Medlar Press.
-
Hewitt E.R. (1948). "A Trout and Salmon Fisherman for Seventy-Five Years". Charles Scribner's Sons.
-
Heschl, Adolf. (1989). Integration of “Innate” and “Learned” Components within the IRME for Mussel Recognition in the European Bitterling Rhodeus amarus (Bloch). Ethology 81, 193-208 (1989). Available online
- Hill J W (1930) "A Summer on the Test"
- Hinde, Robert (1970). "Animal Behaviour: A Synthesis of Ethology and Comparative Psychology". McGraw Hill.
-
Hubálek M. (2020). "A Brief (Hi)Story of Just-So Stories in Evolutionary Science". Philosophy of the Social Sciences. August 2020. Available online
-
Hughes, Dave (1987). "Handbook of Hatches: A basic guide to identifying local foods and selecting flies to match them". Stackpole Books.
-
Hughes, D. Trout Flies: The Tier's Reference. 1999. Stackpole Books.
-
Hughes, Dave (2005)."Handbook of Hatches: Introductory Guide to the Foods Trout Eat and the Most Effective Flies to Match Them". Stackpole Books.
-
Hutchinson, John M.C. , and Gigerenzer, Gerd. Simple heuristics and rules of thumb: Where psychologists and behavioural biologists might meet. Behavioural Processes 69 (2005) 97–124.
Available online
-
Ishii, Yumiko & Shimada, Masakazu. (2009). The effect of learning and search images on predator–prey interactions. Popul Ecol. 52. 27-35. Available online
-
Jacyna, Stephen 2009."The most important of all the organs:Darwin on the brain". Brain 2009: 132; 3481–3487. Available online
-
Johnsson, Jörgen & Kjällman-Eriksson, Karin. (2008). Cryptic prey colouration increases search time in brown trout (Salmo trutta): Effects of learning and body size. Behavioral Ecology and Sociobiology. 62. 1613-1620. 10.1007/s00265-008-0590-8. Available online
-
Juracek, John (2016).
"Does fly pattern matter? How often is fly choice your limiting factor".
Available online
-
Juracek, John (2020). "A Failure Of Modern Fly Design". Available online
-
Juracek, John (2020a). "A Sign Of The Times, Literally". Available online
-
Juracek John, Pat McCabe and Todd Tanner(2020b).In School of Trout. "Essential Dry Flies Video Class" Available online>
-
Kamil, Alan and Bond, Alan B., "Selective Attention, Priming, and Foraging Behavior" (2006). Papers in Behavior and Biological Sciences. Paper 40. Available online
-
Kašpar. Břetislav (2017) "Tup's Indispensable". Global Flyfisher. Published Mar 17. 2017. Available online
- Kenyon, Paul (2020) 'How does a trout catch a fly' Available online
-
Kenyon, Paul (May 2020) "How does a trout see a fly?". The Field, May 30th 2020.
Available online
-
Kettlewell H. B. D. (1959) "Darwin's Missing Evidence: Changes in Moths Observed" reprinted in Science on February 20, 2009. Available online
-
Kloepper (2016) Visual predators find cryptic prey more effectively by learning a search image. Available online
-
Klewein Kent (2020). "6 Easy Tips to Help Fly Anglers Catch Educated Trout". Gink & Gasoline Blog. Posted on June 16, 2020. Available online
- La Branche, George M L (1914) "The Dry Fly and Fast Water" Charles Scriber's Sons, New York. Available online at:
Available online
-
Lafontaine, Gary (1990)"The Dry Fly: New Angles". The Lyons Press.Sample available online
-
Lagogiannis K, Diana G, Meyer MP. (2020). Learning steers the ontogeny of an efficient hunting sequence in zebrafish larvae. Elife. 2020;9:e55119. Published 2020 Aug 10. doi:10.7554/eLife.55119 Available online
-
Lawton, Terry (2005). "Nymph Fishing a History of the Art and Practice". Stackpole Books. Elements available online
-
Lett, Bow Tong (1980). "Taste potentiates color-sickness associations in pigeons and quail". Animal Learning & Behavior 1980,8(2),193-198.
-
Linton. Paul (2020). "Does vision extract absolute distance from vergence?". Attention, Perception, & Psychophysics volume 82, pages 3176–3195(2020), Available online
-
Lorenz, Konard (1981)."The Foundations of Ethology", trans. Konrad Z. Lorenz and Robert Warren Kickert, New York: Springer Science+ Business. Preview available online
-
McCaskie, H.B. (1950). The Guileless Trout. Cresset Press.
-
McDonnell, Paddy. (2016) "The Vulnerable Prey Image (Part 2)". Blog entry March 19, 2016. Available online
-
McDonnell, Paddy. (2016) "The Vulnerable Prey Image (Part 1)". Blog entry March 19, 2016. Available online
-
McFarland David (1985).Animal Behaviour. Pitman.
-
MacKay, Barry (1974).
"Conditioned food aversion produced by toxicosis in atlantic cod",
Behavioral Biology,
Volume 12, Issue 3,
1974,
Pages 347-355.
Abstract available online
-
McNeil, Carl (undated). Interview in "Imitation or Presentation?". Midcurrent. Available online
-
Marinaro, V.C. (1970).A Modern Dry-Fly Code. Nick Lyons Books.
- Martinowksy (2007)
image of moths on tree. Attribution:
By Martinowksy, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=1855638
By Martinowksy, CC BY-SA 3.0
Link to larger version of image
-
Mason, Bill (1994). Fly Fishing: Learn from a Master. Sports Illustrated Books.
-
Mason, Bill (1994). Fly Fishing: Learn from a Master. Sports Illustrated Books.
-
Mirza, Reehan and Chivers, Douglas (2000). Predator-recognition training enhances survival of brook trout: Evidence from laboratory and fielden closure studies. Can. J. Zool.78: 2198–2208 (2000)
- Neuswanger Jason, Mark S. Wipfli, Amanda E. Rosenberger and Nicholas F. Hughes. (2014) Mechanisms of drift-feeding behavior in juvenile Chinook salmon and the role of inedible debris in a clear-water Alaskan stream. Environ Biol Fish (2014) 97:489–503 available online
-
Norris, T. (1865). The American angler's book: embracing the natural history of sporting fish, and the art of taking them. New edition. Philadelphia: E. H. Butler. Available online
-
Nylander, Rolf (2020). "Floating Flies | Full Film 2020". Available online
-
O'Reilly, Pat (2006). "Matching the Hatch: Stillwater, River & Stream". Swan Hill Press.
-
Overfield, (undated)."Skues and The World's Best Rod)". Flyfishers' Journal. p 96-8. Available online
- Pachur, Thorsten , Peter M. Todd, Gerd Gigerenzer, Lael J. Schooler and
Daniel G. Goldstein.(2011)
The recognition heuristic: a review of theory and tests Front. Psychol., 05 July 2011
-
Parrella, Michael P. (2013). "A History of the Entomology of Fly Fishing", American Entomologist, Volume 59, Issue 1, Spring 2013, Pages 15–27, Available online
-
Pearce, John M. (1997). Animal Learning and Cognition. Psychology Press.
-
Pilkington, David (2021) in Arundell Arms Hotel Spring 2021 newsletter,
-
Pinker, Steven (2004) "Why nature & nurture won’t go away". Dædalus Fall 2004: 5-17. Available online
-
Plaisted, K. C., & Mackintosh, N. J. (1995). Visual search for cryptic stimuli in pigeons: Implications for the search image and search rate hypotheses. Animal Behaviour, 50(5), 1219–1232.Available online
-
Proper, Datus.(1993) "What the Trout Said: About the Design of Trout Flies and Other Mysteries". Swan Hill Press.
-
Randall, Jason. "What Aboutt Refusals?", American Angler, undated.Available online
-
Randall, Jason. (2014) Trout Sense . Stackpole / Headwater. Kindle Edition. Stackpole Books.
-
Raveh, Shirley , Kathrin Langen, Theo C. M. Bakker, Christian Josephs, Joachim G. Frommen (2018). Oddity, predation risk and social decisions in aquatic invertebrates Ethology. 2018;1–8.
Available online
-
Reebs, Stéphan G. (2008) Long-term memory in fishes. Université de Moncton, Canada.Available online
-
Reichenthal, A., Segev, R. & Ben-Shahar, O. Feature integration theory in non-humans: Spotlight on the archerfish. Atten Percept Psychophys 82, 752–774 (2020).
Available online https://doi.org/10.3758/s13414-019-01884-4
- Rennie, James (1833) "Alphabet of Scientific Angling".
Available online
- Roberts, J (1994). To Rise a Trout.Marlborough, The Crowood Press.
-
Rollo, W. Keith (1944 ). Fly Fishing: Practical Hints on the Sport and Choice of Tackle and Water. H.F.& G.Witherby Ltd.,
-
Rolston, Tim (2016). "Vulnerability, A super stimulus?". The Fishing Gene blog. March 7th 2016 Available online
-
Rolston, Tim (2013) "The Cuckoo and the Trout". The Fishing Gene blog. August 31st 2013. Available online
-
Rolston, Tim (2020). "It’s NOT about the fly". The Fishing Gene blog. December 13th 2020. Available online
-
Ronalds, Alfred (1836)."The Fly-fisher's Entomology". Available online
-
Royal Entomological Society (2012). "Handbooks for the identification of British insects". Vol 1X Part 1.
Available online
-
Rudge, David W. (2005) The Beauty of Kettlewell's Classic Experimental Demonstration of Natural Selection. BioScience, Volume 55, Issue 4, April 2005, Pages 369–375.
Available online
-
Ruxton, Graeme D. , Tom N. Sherratt, and Michael P. Speed (2007)."Avoiding Attack: The Evolutionary Ecology of Crypsis, Warning Signals and Mimicry" Published to Oxford Scholarship Online: September 2007. Abstract & list of contents available online
-
Schill Daniel J. , J. S. Griffith & Robert E. Gresswell.(1986)" Hooking Mortality of Cutthroat Trout in a Catch-and-Release Segment of the Yellowstone River, Yellowstone National Park".North American Journal of Fisheries Management Volume 6, 1986 - Issue 2 Abstract available online
-
Schooler Lael J , Ralph Hertwig and Stefan M. Herzog (2012).
"How Smart Forgetting Helps Heuristic Inference".
Chapter 6 (p144-166) In
Ecological Rationality: Intelligence in the World.
Peter M. Todd and Gerd Gigerenzer (Eds).
Print publication date: 2012
Print ISBN-13: 9780195315448.
Published to Oxford Scholarship Online: May 2012 Available online
- Schullery, Paul (1987) "American Fly Fishing: A History" Lyons & Burford (p119-120).
-
Schullery, Paul (2002). "History and Mr. Gordon".
The American Fly Fisher, Journal of the American Museum of Fly Fishing Vol 28, No 1, 2-11, 2002.
Available online
-
Schullery, Paul (2007) "Norris on Trout Fishing: A Lifetime of Angling Insights". By Stackpole Books. Limited preview available online
- Schweibert, Ernest (1979) "Trout". Andre Deutch Ltd. London.
-
Schwartz, Barry. (1974). On going back to nature: A review of Seligman and Hager's Biological boundaries of learning. Journal of the Experimental Analysis of Behavior. 21. 10.1901/jeab.1974.21-183. Available online
- Seligman M E P. On the generality of the laws of learning.Psychol. Rev.77:406-18, 1970.
-
Seligman, M. E. P. & Hager, J. L. (1972, August). Biological boundaries of learning. The sauce-bearnaise syndrome. Psychology Today, V6, 59-61, 84-87.
- Seligman M E P. (1980). This Week’s Citation Classic. Current Contents Number 8, February 25, 1980. Available online
-
Shettleworth Sara J. (1974) .
Function, Causation, Evolution, And Development Of Behavior: A Review Of The Animal In Its World, By N.tinbergen.
Journal Of The Experimental Analysis Of Behavior.
1974, 22, 581-590.
Number 3 (November) Available online
-
Siebeck, Ulrike & Litherland, Lenore & Wallis, G. (2009). Shape learning and discrimination in reef fish. The Journal of experimental biology. 212. 2113-9. 10.1242/jeb.028936. Available online
-
Skues, G. E. M. (1921). The Way of a Trout with the Fly: And some further studies in minor tactics. London: Adams and Charles Black. Extract available online
- Skues, G. E. M. (1914). Minor Tactics of the Chalk Stream and Kindred Studies (2nd ed.). London: A & C Black. Available online
-
Smith, Robert (Undated). "Gold Ribbed Hare’s Ear". The Sliding Stream Blog. Available online
-
Steeves III, Harrison R. and Ed Koch. (1994). "Terrestrials: A Modern Approach to Fishing and Tying with Synthetic and Natural Materials". Stackpole Books.
-
Supinski, Matt (2014). Selectivity. Stackpole Books.
-
Sutcliffe, Tom (2013). "What fly rod did GEM Skues rate as his best and why?". Available online
"
-
Swentosky, Domenick (2019)."You don’t have to match the hatch". Troutbitten Blog. Jun 12, 2019. Available online
Swisher, D. & Carl Richards (1975). "Fly Fishing Strategies". Lyons & Burford.
-
Swisher, Doug (2016) Strategies for Selective Trout. YouTube video. Available here
- Swisher, Doug and Carl Richards (2018) "Selective Trout The Last Word on Stream Entomology and Aquatic Insect Imitation".
Kindle Edition
- Smith, Robert "Dr William Baigent".
Available online
-
Tapply, William G. (2016 reissue) “The Truth about Fly Fishermen,” Available online
- Tapply, William G. (2017 reissue)"How to Fool Really “Smart” Trout". Available online
-
The Fly fishers Club Spring Journal 1938 The Nymph Debate. Available on request
-
Todd PM, and Gigerenzer G. (2000). Précis of Simple heuristics that make us smart. Behav Brain Sci. 2000 Oct;23(5):727-41; discussion 742-80. Available online
-
Todd Peter M. and Gerd Gigerenzer (2012)."What Is Ecological Rationality?". Chapter 1 (p3-32) In
Ecological Rationality: Intelligence in the World.
Peter M. Todd and Gerd Gigerenzer (Eds).
Print publication date: 2012
Print ISBN-13: 9780195315448.
Published to Oxford Scholarship Online: May 2012 Available online
-
Travis, Tom (2016). "Skues, wet flies, nymphs and the angler".
Available online
-
Treisman Anne M. & Garry Gelade (1980)
"A feature-integration theory of attention". Cognitive Psychology, Volume 12, Issue 1, 1980, Pages 97-136. Abstract available online
-
Tinbergen, L. (1960). The natural control of insects in pinewoods. 1. Factors influencing the intensity of predation by songbirds. Archives Néerlandaises de Zoologie, 13, 265-343.
-
Trivedi,Chintan A. and Johann H. Bollmann (2013).Visually driven chaining of elementary swim patterns into a goal-directed motor sequence: a virtual reality study of zebrafish prey capture. Neural Circuits, 10 May 2013. Available online
-
von Uexkull, Jacob. (1934)"A stroll through the worlds of animals and men:A picture book of invisible worlds". Monograph available online
-
Ware, Daniel M. (1971a) The predatory behaviour of rainbow trout (Salmo gairdneri). PhD thesis. Department of Zoology, The University of British Columbia Vancouver 8. Canada
-
Weaver, Mike. (1991). "The Pursuit of Wild Trout". Merlin Unwin Books,
- Whitelaw, Ian (2015). 'The History of Fly Fishing in Fifty Flies' Arum Press.
-
Wikipedia (last edited 15 December 2020)."The Fly-fisher's Entomology". Available online
- Wyatt, Bob (2004). "Trout Hunting: The Pursuit of Happiness". Swan Hill Press.
- Wyatt, Bob. (2013). "What Trout Want: The Educated Trout and Other Myths" Stackpole Books.
-
Wyatt, Bob. (undated). "What Trout Want: A Beautiful Fiction". Excerpted from What Trout Want: The Educated Trout and other Fly-fishing Myths. Published in MidCurrant Available online
-
Young, Roger G. and Hayes, John W. (2004). "Angling Pressure and Trout Catchability: Behavioral Observations of Brown Trout in Two New Zealand Backcountry Rivers. North American Journal of Fisheries Management", 2004/11/01.
-
Zupanc, Günther K. H.(2010), Scholarpedia, 5(10):5306. Available online
Link to larger version of image